Resumos
Em áreas úmidas, como o Pantanal, a composição florística e estrutura comunitária respondem principalmente ao pulso de inundação. E para o entendimento das espécies que adentram esse bioma, essas comunidades de plantas precisam ser descritas florística e estruturalmente. O presente estudo teve como objetivos: caracterizar a composição florística e a estrutura da vegetação sobre uma cordilheira na região do Pirizal, Município de Nossa Senhora do Livramento ,MT; definir o tipo de vegetação e determinar se há maior grau de semelhança florística entre a vegetação estudada com outras áreas de cerrado s s. ou de cerradão do Centro-Oeste brasileiro. Foram alocadas 102 parcelas contíguas 20 x 5 m distribuídas sistematicamente em seis transecções dispostas no sentido borda-centro-borda da cordilheira, totalizando 1,02 ha de área amostrada. Mediram-se os indivíduos com DAP >3 cm, incluindo mortos e palmeiras. Foram identificadas 82 espécies arbustivo-arbóreas, distribuídas em 63 gêneros, 32 famílias e uma espécie indeterminada. Sendo 3 espécies características de áreas com cerradão; 19 do cerrado s.s., nove distribuídas tanto em áreas de cerradão quanto em cerrado s.s., e 10% que pertencem a ambientes inundáveis no Pantanal. Comparando a área de estudada com outras comunidades através da DCA, os resultados indicaram a existência de dois grupos florísticos relativamente distintos, o primeiro formado por áreas compostas por cerrado s.s. e o segundo por áreas com cerradão. Entre as áreas formadas por cerradões, ocorreu uma separação das áreas amostradas na região do no Pantanal (áreas inundáveis) daquelas que ocorrem em outras localidades do Centro-Oeste (áreas secas). A vegetação estudada sobre cordilheira apresentou maior relação florística e estrutural com os cerradões do que com os cerrados s.s. do Centro-Oeste, podendo, dessa forma, ser caracterizada como uma fitofisionomia de cerradão. Os cerrados do Pantanal apresentaram menor relação florística com os de outras regiões do Centro-Oeste, essa heterogeneidade florística pode estar relacionada ao pulso de inundação que atua como filtro para a maioria das espécies das outras áreas do Planalto Central.
comparação florística; cordilheira; cerradão; cerrado sensu stricto
In wetlands as the Pantanal, the floristic composition and community structure respond primarily to the flood pulse and, in order to understand the species that enter this biome, these plant communities need to be floristically and structurally described. Therefore, the objectives of this paper are characterize floristic composition and vegetation structure on a cordilheira (paleolevees), in the Pirizal region, Nossa Senhora do Livramento County, MT; define the type of vegetation and determine whether there is a greater floristic similarity degree between the object of this study and other cerrado s.s. or cerradão areas in Central Brazil. 102 contiguous plots of 20 x 5 m were distributed systematically arranged in six transects towards edge-center-edge cordilheira, thus having a total area of 1,02 ha. For sampling purposes, only individuals with entire stem DBH > 3 cm including dead trees and palms. 82 different species of trees and shrubs were identified, which in turn were spread in 63 genera, 32 families and one unidentified species. Being 37 species characteristics of areas with cerradão, 19 of cerrado s.s., nine distributed both in areas of cerradão and in cerrado s.s., and 10% belonging to the Pantanal wetland environments. Comparing the studied area with other communities across the DCA, the results indicated the existence of two floristic groups relatively distinct, the first formed by areas composed of cerrado s.s. and the second by cerradões areas. Among the areas formed by cerradões, there was a separation of the sampled areas in the region of the Pantanal (wetlands) those occurring elsewhere in the Centro-Oeste (dry land). The vegetation on cordilheira showed greater floristic and structural relationship with cerradões than with cerrado s.s. of west center region of Brazil, and may thus be characterized as a vegetation type of cerradão. The savannas of the Pantanal had a lower floristic relationship with other regions of the Centro-Oeste, this floristic heterogeneity may be related to the flood pulse which acts as a filter for most species in other areas of the Planalto Central.
florist comparasion; cordilheira; cerradão; cerado sensu stricto
ARTIGOS
Caracterização da flora e estrutura do estrato arbustivo-arbóreo de um cerrado no Pantanal de Poconé, MT
Characterization of the flora and structure of the shrub-tree layer in a cerrado area in Pantanal do Poconé, MT
Cândida Pereira da CostaI, * * Autor para correspondência: Cândida Pereira da Costa, e-mail: candidapcosta@gmail.com ; Cátia Nunes da CunhaII; Silmara Cardoso CostaII
IPrograma de pós-Graduação em Ciências Florestais, Laboratório de Manejo Florestal, Universidade de Brasília - UnB, CP 04357, CEP 70919-980, Campus Universitário Darcy Ribeiro, Brasília, DF, Brasil
IIDepartamento de Botânica e Ecologia, Instituto de Biociências, Universidade Federal de Mato Grosso - UFMT, Av. Fernando Corrêa da Costa, s/n, CEP 78060-900, Cuiabá, MT, Brasil, e-mail: catianc@ufmt.br
ABSTRACT
In wetlands as the Pantanal, the floristic composition and community structure respond primarily to the flood pulse and, in order to understand the species that enter this biome, these plant communities need to be floristically and structurally described. Therefore, the objectives of this paper are characterize floristic composition and vegetation structure on a cordilheira (paleolevees), in the Pirizal region, Nossa Senhora do Livramento County, MT; define the type of vegetation and determine whether there is a greater floristic similarity degree between the object of this study and other cerrado s.s. or cerradão areas in Central Brazil. 102 contiguous plots of 20 x 5 m were distributed systematically arranged in six transects towards edge-center-edge cordilheira, thus having a total area of 1,02 ha. For sampling purposes, only individuals with entire stem DBH > 3 cm including dead trees and palms. 82 different species of trees and shrubs were identified, which in turn were spread in 63 genera, 32 families and one unidentified species. Being 37 species characteristics of areas with cerradão, 19 of cerrado s.s., nine distributed both in areas of cerradão and in cerrado s.s., and 10% belonging to the Pantanal wetland environments. Comparing the studied area with other communities across the DCA, the results indicated the existence of two floristic groups relatively distinct, the first formed by areas composed of cerrado s.s. and the second by cerradões areas. Among the areas formed by cerradões, there was a separation of the sampled areas in the region of the Pantanal (wetlands) those occurring elsewhere in the Centro-Oeste (dry land). The vegetation on cordilheira showed greater floristic and structural relationship with cerradões than with cerrado s.s. of west center region of Brazil, and may thus be characterized as a vegetation type of cerradão. The savannas of the Pantanal had a lower floristic relationship with other regions of the Centro-Oeste, this floristic heterogeneity may be related to the flood pulse which acts as a filter for most species in other areas of the Planalto Central.
Keywords: florist comparasion, cordilheira, cerradão, cerado sensu stricto.
RESUMO
Em áreas úmidas, como o Pantanal, a composição florística e estrutura comunitária respondem principalmente ao pulso de inundação. E para o entendimento das espécies que adentram esse bioma, essas comunidades de plantas precisam ser descritas florística e estruturalmente. O presente estudo teve como objetivos: caracterizar a composição florística e a estrutura da vegetação sobre uma cordilheira na região do Pirizal, Município de Nossa Senhora do Livramento ,MT; definir o tipo de vegetação e determinar se há maior grau de semelhança florística entre a vegetação estudada com outras áreas de cerrado s s. ou de cerradão do Centro-Oeste brasileiro. Foram alocadas 102 parcelas contíguas 20 × 5 m distribuídas sistematicamente em seis transecções dispostas no sentido borda-centro-borda da cordilheira, totalizando 1,02 ha de área amostrada. Mediram-se os indivíduos com DAP >3 cm, incluindo mortos e palmeiras. Foram identificadas 82 espécies arbustivo-arbóreas, distribuídas em 63 gêneros, 32 famílias e uma espécie indeterminada. Sendo 3 espécies características de áreas com cerradão; 19 do cerrado s.s., nove distribuídas tanto em áreas de cerradão quanto em cerrado s.s., e 10% que pertencem a ambientes inundáveis no Pantanal. Comparando a área de estudada com outras comunidades através da DCA, os resultados indicaram a existência de dois grupos florísticos relativamente distintos, o primeiro formado por áreas compostas por cerrado s.s. e o segundo por áreas com cerradão. Entre as áreas formadas por cerradões, ocorreu uma separação das áreas amostradas na região do no Pantanal (áreas inundáveis) daquelas que ocorrem em outras localidades do Centro-Oeste (áreas secas). A vegetação estudada sobre cordilheira apresentou maior relação florística e estrutural com os cerradões do que com os cerrados s.s. do Centro-Oeste, podendo, dessa forma, ser caracterizada como uma fitofisionomia de cerradão. Os cerrados do Pantanal apresentaram menor relação florística com os de outras regiões do Centro-Oeste, essa heterogeneidade florística pode estar relacionada ao pulso de inundação que atua como filtro para a maioria das espécies das outras áreas do Planalto Central.
Palavras-chave: comparação florística, cordilheira, cerradão, cerrado sensu stricto.
Introdução
No Pantanal de Mato Grosso diferentes regiões fitogeográficas se interpenetram, ocorrendo espécies vinculadas ao Cerrado; à Floresta Tropical Úmida e a Seca e Chaco, Prance & Schlaller (1982), Nunes da Cunha (1999, 2001), Pott, A. & Pott, V.J. (1994, 1999). Segundo Silva et al. (2000) a vegetação savânica (cerrado) ocupa cerca de 70% do total dos tipos de vegetação no Pantanal. De acordo com Eiten (1982) e Sarmiento (1984) o Pantanal é considerado uma savana hipersazonal o que significa uma vegetação savânica sujeita a dois estresses contrastantes, um induzido pela seca e outro pela prolongada inundação.
Na comparação florística-geográfica de Castro (1994), os cerrados do Pantanal, principalmente os localizados em suas partes Nordeste, Leste e Sudeste, são contínuos aos do Planalto Central, pois a maioria das espécies encontradas naquela área úmida são as mesmas do Planalto. As comparações floristicas da comunidade arbórea do Cerrado realizada por Ratter et al. (2003) em 376 sítios de Cerrado foram correspondentes as de Castro, classificando o Pantanal no grupo fitogeográfico Centro-Oeste.
Grande parte das espécies lenhosas no Pantanal, não é exclusiva de região fitogeográfica do Cerrado, o que significa que no Pantanal ocorrem outros grupos de plantas de outras diferentes províncias fitogeográficas tais como Floresta Estacional, Chaco e da Bacia Amazônia (Nunes da Cunha & Junk 1999, 2001).
A maioria das 116 espécies lenhosas dominantes da flora do cerrado brasileiro arroladas por Ratter et al. (2006) e muitas das árvores mais frequentes das florestas estacionais (Jardim et al. 2003, Oliveira-Filho et al. 2006) também ocorrem no Pantanal em floresta transicionais ou em comunidades separadas.
Na fisionomia pantaneira, o cerradão ocupa áreas mais elevadas (cordilheiras = paleodiques) e o cerrado sensu stricto, nas áreas mais baixas, tendendo para campo à medida que aumenta o grau de inundação (Silva et al. 2000, Nunes da Cunha & Junk 2001). Em anos excepcionais, as cordilheiras podem inundar, e isso reflete diretamente na biota, que responde com composição e estrutura de comunidade especifica (Junk et al. 1989).
Aplicando o conceito de Forman & Godron (1986), em relação à paches na paisagem, o Pantanal constitui um grande mosaico de vegetação onde os retalhos são definidos pelas fisionomias, composição florística e tipos de vegetações, este mesmo padrão foi considerado por Borges & Shepherd (2005) para o Cerrado. O entendimento dos patches (retalhos) implica no conhecimento da vegetação, processos geomorfológicos, regimes de perturbação e interferência humana, o que definirá suas características (Forman & Godron 1986). O primeiro passo nesse sentido está na determinação da composição de espécies e da forma como as mesmas estruturam a comunidade no espaço. Estudos florísticos e fitossociológicos foram desenvolvidos em vegetação de cerrado em diferentes regiões do país (Ribeiro et al. 1985, Nascimento & Saddi 1992, Felfili et al. 1993, Ratter et al. 1996, Felfili et al. 2000, 2002, 2007, Assunção & Felfili 2004, Teixeira et al. 2004, Balduino et al. 2005, Barbosa et al. 2005, Marimon Jr. & Haridasan 2005, Gottsberger & Silberbauer-Gottsberger 2006, Batalha & Martins 2007, Neri et al. 2007, Ishara et al. 2008, Sasaki & Mello-Silva 2008). Grande parte dos trabalhos acima citados descreve gradientes e/ou patches da vegetação do Cerrado.
No Pantanal, as pesquisas fitossociológicos como as Ratter et al. (1988, 2003), Borges & Shepherd (2005), Pott, A. & Pott, V.J. (1994), Prado et al. (1994), Dubs (1994), Guarim-Neto et al. (1996), Schessl (1999), Nunes da Cunha & Junk (1999), Pott, A. & Pott, V.J. (1999), Salis et al. (1999), Silva et al. (2000), Nunes da Cunha (2001, 2004), Damasceno-Jr. et al. (2004, 2005), ainda não são suficientes para apontar a significativa riqueza florística e as diferenças estruturais e sua relações com o ambiente físico.
Considerando ainda, que a vegetação sobre cordilheiras apresenta diferentes tipologias vegetacionais, sendo hábitats-chave para fauna e flora e locais onde processos ecológicos fundamentais tais como, produção de serapilheira e ciclagem de nutrientes tendo efeito em toda a Zona de Transição Aquática Terrestre (ATTZ), Junk et al. (1989). Por isso estas comunidades ainda carecem serem descritas quanto a sua composição e estrutura fitossociológica, pois medidas de abundância e de distribuição das espécies são importantes, quando se tem o objetivo de desenvolver uma base teórica que subsidie seu manejo, conservação ou a recuperação de áreas similares.
Para entender o Pantanal como parte do bioma Cerrado e/ou o Pantanal como um bioma, comparações florísticas que descrevam o padrão de distribuição geográfico da flora entre essas regiões é o primeiro passo nesse sentido.
Diante do exposto, espera-se que a vegetação estudada tenha maior semelhança florística com áreas de cerradões do que com os cerrados s.s.; além disso, os cerrados do Pantanal podem ser considerados como uma unidade diferenciada dentro da região do Centro-Oeste. Este fato se deve a saturação hídrica que atua como filtro no estabelecimento das espécies lenhosas. Para atender a estes pressupostos esse trabalho teve como objetivos, caracterizar a composição florística e a estrutura da vegetação sobre cordilheira; definir o tipo de vegetação e determinar se há maior grau de semelhança florística entre a vegetação estudada com outras áreas de cerrado sensu stricto ou de cerradão do Centro-Oeste brasileiro.
Material e Métodos
1. Localização e descrição geral da área de estudo
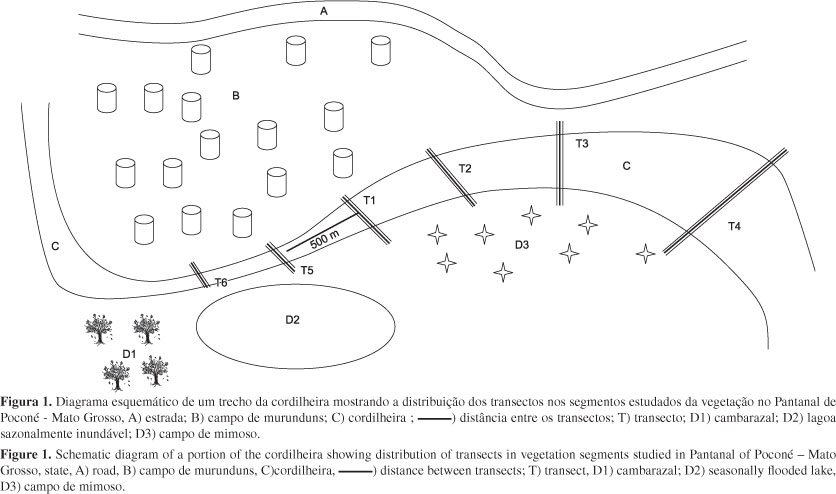
A vegetação estudada localiza-se sobre uma cordilheira (cordão arenoso livre de inundações anuais) da Fazenda Nossa Senhora da Aparecida na região denominada Pirizal situada a Nordeste do Pantanal de Poconé (16° 15' 12'' S e 56° 22' 12'' W), município de Nossa Senhora do Livramento, Mato Grosso. Localizando-se ainda e entre uma depressão inundável e campo de murunduns, o alagamento aconteça somente no habitas de entorno. A vegetação é formada por um estrato lenhoso arbóreo-arbustivo dominante, e por um estrato herbáceo com presença de Bromelia balansae Mez. A área de estudo e as fitofisionomias adjacentes passaram por incêndios e desmatamento no ano de 1999, cerca de um ano antes do inicio deste trabalho.
O clima é do tipo AW de acordo com classificação de Köppen com duas estações bem definidas: a estação seca, de maio para setembro e a estação chuvosa, de outubro para abril (Nunes da Cunha & Junk 2004). A precipitação máxima é 1.384 mm, no mês de janeiro com precipitação mínima no mês de julho. A temperatura média anual é de 25,8 °C, sendo outubro o mês mais quente, enquanto julho é o mais frio (Rebellato & Nunes da Cunha 2005). No Pantanal duas estações são observadas durante o ano, sendo uma seca, no período de maio a setembro e a outra chuvosa, de outubro a abril (Nunes da Cunha & Junk 2004). O processo de inundação sazonal é dividido em quatro fases: enchente, cheia, vazante e seca (Rebellato & Nunes da Cunha 2005).
2. Amostragem e análise dos dados
O levantamento florístico consistiu na coleta mensal de material botânico das espécies arbustivo-arbóreas ao longo de um ano (2000-2001). As espécies foram identificadas por especialistas e por comparações com amostras de herbário. O material coletado foi herborizado e depositado no Herbário Central da Universidade Federal de Mato Grosso - UFMT. As espécies foram classificadas em famílias de acordo com o sistema do Angiosperm Phylogeny Group (2009). Através do material coletado durante os levantamentos florísticos foi elaborada uma listagem contendo as famílias, gêneros e espécies. Os nomes das espécies e de seus autores foram conferidos através de consultas ao W3 Trópicos (Missouri Botanical Garden, 2010).
Para as amostragens fitossociológicas foram alocadas 102 parcelas contíguas 20 × 5 m, totalizando uma área amostral de 1,02 ha. As parcelas foram distribuídas sistematicamente em seis transecções dispostas no sentido borda-centro-borda da cordilheira, para captação das mudanças no gradiente ambiental (Figura 1).
Quanto à forma de vida, foram considerados como hábito arbóreo os indivíduos com caule indiviso, altura >4 m e diâmetro >5 cm; como arbustivos aqueles com caule indiviso ou ramificado desde a base, altura variando de 0,5 a 4 m e diâmetro >1 cm (Rizzini 1997).
Para a analise estrutural, as medidas do diâmetro do caule foram tomadas a 1,30 do solo nos indivíduos que apresentaram DAP > 3 cm, inclusive para indivíduos mortos e palmeiras, excluindo da amostragem apenas lianas. As medidas de altura dos indivíduos foram estimadas com vara graduada.
Na análise estrutural foram calculados os valores absolutos e relativos de densidade, frequência e dominância (Muller-Dombois & Ellemberg 1974). A soma desses parâmetros, em termos relativos, definiu o Valor de Importância (VI) das espécies. Para as famílias calculou-se o Índice de Valor de Importância Familiar (FIV) de acordo com Mori et al. (1983), onde FIV = (Diversidade Relativa + Abundância Relativa + Dominância Relativa) × 100. Além disso, para obter melhor entendimento da estrutura da vegetação na área, foi elaborado um histograma de frequência de classes de diâmetro e de altura para todos os indivíduos da área. O número de classes diamétricas (K) foi obtido mediante cálculos matemáticos pelo modelo de Sturges (1926): K = 1+3,3Ilogn em que: n = número de elementos da amostra. E o intervalo de classes (h): h = H/K em que: H = amplitude total (DAPmáximo - DAPmínimo). A distribuição das alturas foi realizada com intervalos de classe de 1,0 m.
As diversidades florísticas foram avaliadas através dos índices Shannon-Wiener e o α de Fisher (Magurran 1988). O primeiro índice foi obtido pela fórmula: H' = - Σ (pi) (logn pi), onde pi = (ni / N) é a probabilidade de que um indivíduo amostrado pertença a espécie i; ni = no total de indivíduos da espécie i; N = no total de indivíduos amostrados na área; a equitabilidade (uniformidade) foi obtida segundo a fórmula E' = H' /logn S, onde S = número de espécies. E o segundo (α de Fisher) foi calculado para este trabalho, e outros desenvolvidos em áreas de cerrado sensu stricto e cerradão, que forneceram o número de indivíduos amostrados e de espécies identificadas com diâmetro mínimo de inclusão >30 cm (Diâmetro a Altura do Solo, DAS); 3 e 5 cm de Diâmetro a Altura do Peito (DAP), Tabela 1. Sendo que este último índice é recomendado para comparações da diversidade em áreas com tamanho amostral diferenciado, pois ele é insensível ao tamanho da área amostral (Magurran 1988).
Para avaliar as relações florísticas com os resultados desse estudo, foram selecionadas outras 28 áreas (sendo 13 cerrado s.s. e 15 cerradões), Tabela 1. As 28 listas de espécies foram convertidas em uma matriz de presença-ausência com 221 espécies (sendo incluído apenas espécies arbustivas, arbóreas e palmeiras). Para não ocorrer o falso aumento das espécies e a falsa diferença florística entre as áreas devido à incorreção no uso de nomes, a sinonímia e a grafia dos taxa foi conferida consultando o endereço eletrônico da Missouri Botany Garden (http: www.mobot.org.) e do The Plant Names Project 2000). Em seguida, fez-se uma ordenação das 28 áreas, por meio de uma análise de correspondência segmentada (DCA, Detrendend Correspondence Analysis) (Hill & Gauch 1980). Para isso, foi utilizado o programa Pc-Ord for Windows versão 4.0 (McCune & Mefford 1999).
Resultados
Foram identificadas 82 espécies arbustivo-arbóreas, distribuídas em 63 gêneros, 32 famílias e uma espécie indeterminada (Tabela 2). Quanto ao hábito de crescimento das espécies amostradas, 31 (37,80%) foram arbustivas, 50 (61%) arbóreas, incluindo uma (1,22%) palmeira. As famílias que mais contribuíram para a riqueza de espécies foram: Fabaceae (17 espécies), Myrtaceae, Malpighiaceae e Rubiaceae (cinco espécies), Vochysiaceae e Melastomataceae (quatro espécies), Anacardiaceae, Apocynaceae, Bignoniaceae, Combretaceae e Sapindaceae (3 espécies). Estas famílias abrangeram 55 espécies, constituindo 67,1% das espécies amostradas. As 26 espécies restantes estão distribuídas em 21 famílias, das quais 16 foram representadas por uma espécie. Os gêneros com maior riqueza de espécies foram Byrsonima e Eugenia (quatro espécies) seguidos por Alibertia, Qualea e Tabebuia (três espécies).
No levantamento florístico foram encontradas 37 (46,3%) espécies características de áreas com cerradão (Oliveira Filho & Martins 1986, Ratter et al. 1996, 2003), 20 espécies (25%) características do cerrado sensu stricto (Oliveira Filho & Martins 1986, Gottsberger & Silberbauer-Gottsberger 2006), nove espécies (11,3%) distribuídas tanto em áreas de cerradão quanto em cerrado s.s. (Oliveira Filho & Martins 1986, Ratter et al. 1996, 2003), e 10% (8/80) que pertencem a ambientes inundáveis no Pantanal (Damasceno Jr. et al. 1999, Nunes da Cunha & Junk 1999), ocorrendo ainda três espécies (3,75%) que não foram registradas em outras áreas, apenas na área de estudo (Tabela 1).
As medidas de diversidade de Shannon-Wiener e equabilidade de Pielou (H' = 4,72 e E' = 0,76) demonstraram que a área estudada é mais diversa que outras comunidades (Tabela 1). Os valores de α de Fisher entre as áreas comparadas variaram de 4,64 (N = 1009 e S = 25, para o cerradão A7) a 24,53 (N = 1019 e S = 92, para o cerrado s.s. da A25), os maiores valores foram representados para as áreas formadas por cerrado s.s., enquanto os cerradões, principalmente os do Pantanal, apresentaram menores valores. Para a área estudada a alfa-diversidade foi de 17,05 (Tabela 1).
Em 18,6% (13 espécies) registraram-se com somente um indivíduo no levantamento, sendo consideradas raras: Agonandra brasiliensis, Alibertia sp., Brosimum gaudichaudii, Casearia silvestris, Combretum lanceolatum, Diospyros hispida, Dipteryx alata, Erythroxylum suberosum, Psidium guineense, Qualea parviflora, Strychnos pseudoquina, Tabebuia insignis e Tocoyena formosa.
Quanto à frequência as espécies Curatella americana, Magonia pubescens, Hymenaea stigonocarpa, Alibertia edulis, Astronium fraxinofolim apresentaram os valores mais altos, indicando que as mesmas foram registradas em boa parte das parcelas (20 a 50%). A dominância relativa foi significativa para Curatella americana e Magonia pubescens, enquanto as demais espécies apresentaram menores índices.
O número de classes diamétricas encontradas em razão da amplitude dos dados foram 11 com intervalo de 1,63 cm em cada classe. A distribuição dos indivíduos por diâmetro, na área de estudo, tende a ser caracterizada por uma curva do tipo J invertido. Pois apresentou a maior concentração (99%) nos primeiros intervalos das menores classes diamétricas (1,0 a 9,19 cm), considerados indivíduos jovens na comunidade estudada (Figura 2). Observando ainda que as espécies arbustivas de DAP menores, como A. edulis e B. orbignyana também caracterizam o acúmulo nas primeiras classes, pois são frequentes nessa comunidade vegetal (Tabela 3). Na Figura 3 pode-se observar que a maioria dos indivíduos (58%) está no primeiro estrato, entre 1 a 5 m de altura. O segundo estrato, indivíduos com altura média de 5,1 a 10 m, alcançou 37% da densidade total e o terceiro estrato com as plantas emergentes entre 10,1 a 16 m corresponde a 4% da densidade total amostrada.
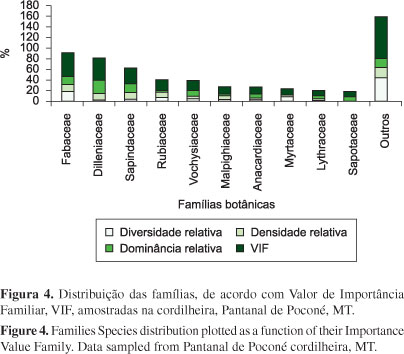
As 10 famílias com maiores valores para o Índice de Importância Familiar (VIF) foram: Fabaceae (VIF = 45,8%), Dilleniaceae (VIF = 40,7%), Sapindaceae (VIF = 32,2%), Rubiaceae (VIF = 20,7%), Vochysiaceae (VIF = 20,4%), Malpighiaceae (VIF = 14%), Anacardiaceae (VIF = 13,6%), Myrtaceae (VIF = 12,3%), Lythraceae (VIF = 10,3%) e Sapotaceae (VI = 9,4%). Essas famílias responderam por 219,4% de um total máximo acumulado de 300% de VIF. Observou-se que nem sempre as famílias com maior diversidade de espécies são as de maiores importância na comunidade (Figura 4). Dilleniaceae, Lythraceae e Sapotaceae com apenas 1,40% ficaram entre as 10 famílias em maior importância sociológica. Enquanto outras sete famílias que apresentaram maior diversidade relativa, como Combretaceae e Apocynaceae (4%), foram consideradas com menor importância ecológica em relação as 33 famílias botânicas analisadas.
A análise de correspondência segmentada, DCA, (Figura 5) expressou uma mudança significativa na composição das espécies entre as s fitofisionomias. No primeiro eixo da ordenação houve uma separação entre as áreas formadas por cerrados s.s. (agrupamento florístico 1) das áreas de cerradão (agrupamento florístico 2). Como exceções, o agrupamento florístico 1 (cerrado s.s.) apresentou ainda duas áreas de cerradões (A5, Nova Xavantina (MT), e A17, Planaltina (DF)). O eixo 1 apresentou um gradiente longo (3,45 S.D.) refletindo uma gradativa mudança na composição florística entre as áreas. Esta análise evidenciou que as áreas de cerradão que ocorrem no Pantanal formam um grupo distinto das outras áreas do cerrado do Brasil Central, exceto duas áreas do Cerrado de Goiás (A27 e A28). Ou seja, diferenciou os cerradões que ocorrem em áreas secas para àqueles que ocorrem em áreas inundáveis. O elevado valor do eixo 1 (0,54) indica que parte substancial da variação total (37%) da composição florística entre as áreas foi expressa ao longo do seu eixo. E a variação restante foi refletida pelo segundo eixo que apresentou um baixo autovalor (0,23).
Discussão
O levantamento florístico apresentou representativa riqueza das espécies arbustivo-arbóreas na área estudada (82 espécies), porém o número de espécies foi influenciado diretamente pelo hábito arbóreo. A menor proporção de espécies arbustivas encontradas na vegetação da cordilheira reforça a discussão que a definição do hábito arbustivo na vegetação do cerrado muitas vezes é subjetiva (Castro et al. 1999). Algumas espécies citadas como herbáceas, por Borges & Shepherd (2005), para uma comunidade de Cerrado no município de Santo Antônio do Leverger, MT, em outros estudos foram descritas como arbustivas. Esses dados reforçam a discussão de Castro et al. (1999), Gottsberger & Silberbauer-Gottsberger (2006), sobre a dificuldade de se definir uma espécie "lenhosa" no cerrado. De acordo com esses autores, tal dificuldade seria em decorrência da ausência de critério que possa ser usado universalmente, ou porque pesquisadores diferem no conceito que utilizam e porque algumas espécies exibem variações no hábito de crescimento entre localidades, desde herbáceo até arbóreo (Borges & Shepherd 2005). Além disso, o desenvolvimento das espécies savânicas do Pantanal, são afetadas pela inundação, fator que dificulta na maioria das vezes a definição sobre o seu hábito de crescimento (Nunes da Cunha & Junk 1999, Junk et al. 1989).
As famílias que descrevem melhor a flora da área estudada também foram, nas observações de Rizzini (1997), as famílias mais importantes na composição da vegetação do cerrado. Goodland (1970) destaca ainda para o cerrado a família Euphorbiaceae (14), também encontrada na comunidade estudada. Já Felfili et al. (2001) destacam além de várias outras famílias importantes para o bioma Cerrado, a família Combretaceae com 16 espécies.
Os gêneros (Byrsonima, Eugenia, Alibertia, Qualea e Tabebuia) que mais contribuíram com a riqueza específica na área estudada são do mesmo modo importantes no cerrado. Goodland (1970) e Heringer et al. (1977) relacionam Annona, Alibertia, Byrsonima, Eugenia, Heteropteris e Qualea entre os gêneros que caracterizam esse tipo de vegetação.
A maioria das espécies encontradas neste estudo caracteriza a vegetação de cerradão, estas espécies foram citadas por Oliveira-Filho & Martins (1986), Ratter et al. (1996, 2003) e Guarim et al. (2000) para a mesma fitofisionomia em localidades distintas. Entretanto, os resultados obtidos no levantamento florístico demonstram expressiva riqueza de espécies e de gêneros, e presença de táxons que caracterizam a flora de cerradão e também do cerrado s.s., indicando que a área estudada serve como exemplo de heterogeneidade da vegetação do cerrado (Borges & Sheperd 2005). Fato que evidencia que as espécies não estão distribuídas uniformemente no bioma Cerrado, devido grande parte destas apresentarem características morfológicas e adaptações fisiológicas às condições climáticas e edáficas que prevalecem na região, reforçando o padrão discutido por Felfili & Silva Jr. (1993), Ratter et al. (1996, 1997, 2003), e Silva et al. (2006).
A maioria dos trabalhos aqui utilizados para efeito de comparação utilizou apenas o índice de Shannon-Wiener para caracterizar a diversidade florística, mas para González-Rivas et al. (2006), o índice de Fisher é a medida de diversidade mais recomendada para comparar a alfa diversidade. O índice de Shannon-Wiener para todos os tipos de vegetação e estratos do bioma Cerrado normalmente varia de 3,5 a 4 nats.ind-1. Na área estudada esse valor alcançou 4, 72 nats.ind-1; corroborando com os dados analisados por Castro (1994) e Castro et al. (1999), Ratter et al. (1996, 2003), Borges & Sheperd (2005), que apontam Mato Grosso com um dos estados brasileiros onde a vegetação do cerrado exibe maior diversidade. A equitabilidade estimada de 0,76 sugere moderada uniformidade nas proporções indivíduos-espécies dentro da comunidade vegetal da cordilheira, de acordo com Uhl & Murphy (1981) a equitabilidade é diretamente proporcional à diversidade e, antagônico à dominância.
Por outro lado, a analise dos valores de α de Fisher demonstrou que a área de estudo apresentou diversidade moderada em relação aos cerrados s.s. da Fazenda Água Limpa, DF (Felfili et al. 2000), Alto Paraíso de Goiás, Vila Propício, Serra da Mesa e Serra Negra, em Goiás (Felfili et al. 2007). Esses resultados expressam que os cerrados s.s. apresentaram maior riqueza específica que os cerradões, pois segundo Fisher et al. (1943), a α diversidade é elevada quando o número de espécies é alto em relação ao número de indivíduos e baixa quando o número de espécies é pequeno em relação ao número de indivíduos. A elevada riqueza florística de espécies dos cerrados s.s. em relação aos cerradões, principalmente os do Pantanal, pode ser explicada pela distribuição espacial das espécies. Nessa região a inundação origina mosaicos em pequenas escalas, padrão diferenciado do observado na área core do cerrado, onde os tipos estruturais são normalmente organizados ao longo de gradientes ambientais (Ab'Saber 1983, Eiten 1994, Ribeiro & Tabarelli 2002)
O estudo fitossociológico permitiu caracterizar a estrutura do estrato arbustivo-arbóreo através da estimativa de parâmetros relacionados com as populações das espécies. Na área estudada, a frequência e dominância de Curatella americana foram determinantes para determinar seu VI, enquanto para Magonia pubescens a densidade e também frequência os parâmetros mais importantes. Salis et al. (1999), listaram Curatella americana em 1° lugar na lista de VI numa savana parque em Mato Grosso do Sul. Segundo Oliveira-Filho (1984) Curatella americana é frequente ainda em outros cerrados da Baixada Cuiabana e Pantanal, em seu estudo esta espécie ocorre quase exclusivamente na faixa limítrofe entre cerrado e vereda. Nunes da Cunha & Junk (1999) citam Curatella americana para o grupo das espécies de ampla distribuição no gradiente de inundação, sendo predominante nos campos de murunduns, por resistirem a curtos períodos de inundação. Entre as espécies encontradas em outras localidades Magonia pubescens é a 1ª colocada na tabela de VI nos cerrados de cordilheira, ela é frequente em cerradão de solo fértil, capão, solos arenosos ou siltosos, com distribuição no CE até MG, PI, GO, MT e MS, Paraguai e Bolívia (Pott, A. & Pott, V.J. 1994). Em cerradão mesotrófico "Carvoeiral" no Pantanal, Guarim et al. (2002) citaram como a 2ª colocada em VI. Segundo Ratter (1971), esta espécie é indicadora de cerradões associados a solos mesotróficos.
Na área de estudo foi observado que os maiores valores de VI (> 10,0) estavam restritos a um grupo de sete espécies. Essas espécies podem ser consideradas as que tiveram maior sucesso na exploração dos recursos ambientais. As demais espécies apresentaram VI pouco significativo, isso é indicativo que predominam para estas espécies baixos valores de densidade, frequência e dominância. Entre estas estão as espécies raras (13, 18,6%) que são espécies que apresentaram apenas um (1) indivíduo (Martins 1991). A consideração sobre a raridade em cordilheiras deve ser tratada de maneira específica uma vez que as fronteiras presentes mostram mudanças abruptas favorecendo a presença de determinadas espécies ao longo do gradiente que podem ser frequentes nas áreas adjacentes. Pagano (1995) descreveu em seu trabalho sobre composição florística e estrutura fitossociológica de uma floresta mesófila semidecídua no Estado de São Paulo que é comum em qualquer estudo fitossociológico à ocorrência de número elevado de espécies representadas por um ou poucos indivíduos dentro da área amostral. Para Martins (1991), isto não significa que tais espécies sejam raras, mas que a maior parte das vezes, apresenta distribuição espacial que determina a ocorrência de poucos indivíduos por unidade de área.
Em termos estruturais da comunidade estudada, as famílias Fabaceae e Dilleniaceae apresentaram maior importância sociológica, visto que Dilleniaceae apresentou menor valor de diversidade relativa em relação a outras famílias, e foi considerada a segunda em importância sociológica. Demonstrando que além da riqueza específica, os parâmetros densidade e dominâncias relativas são fundamentais para avaliar a flora da região.
A distribuição diamétrica da comunidade arbustivo-arbórea da área estudada tende a ser caracterizada por uma curva do tipo J invertido, visto que a maior concentração de indivíduos ocorreu nas primeiras classes, segundo Scolforo (1998) a menor classe diamétrica nos histogramas de distribuição geralmente apresenta a maior frequência de indivíduos. Em áreas de Cerrado, a maior concentração dos indivíduos nas primeiras classes diamétricas pode ser explicada pela própria potencialidade genética da maioria das espécies em apresentarem pequeno porte, ou ainda, diante da possibilidade de abate seletivo, fogo, pisoteio (Felfili & Silva Jr. 1998). A distribuição dos indivíduos por altura indicou a existência de um denso sub-bosque, já que a maior concentração de indivíduos ocorreu nas primeiras classes de altura (1-5 m). Em seguida observou-se um segundo estrato arbóreo (5,1-10 m), onde surgem alguns indivíduos emergentes de até 16 m. Indicando que o cerrado da cordilheira apresentou maior semelhança estrutural com as fitofisionomias de cerradão do que com as de cerrado s.s., já que ocorrem para esta comunidade indivíduos com altura maior que 7 m (25%). De acordo com Gottsberger & Silberbauer-Gottsberger (2006), o cerrado sensu stricto é dominado por árvores e arbustos com dossel geralmente menor que 7 m e o cerradão apresentam árvores maiores que 7 m de altura.
Comparando a área de estudada com outras comunidades através da DCA, os resultados indicaram a existência de um gradiente longo no primeiro eixo, ou seja, a maior parte das espécies se agrupa em setores particulares e somente uma pequena porção das espécies está distribuída por todo o gradiente (Teer Braak 1995). Isso sugere que as 28 áreas formam dois grupos florísticos relativamente distintos, o primeiro formado por áreas compostas por cerrado s.s. e o segundo por áreas com cerradão. Entre as áreas formadas por cerradões, ocorreu uma separação das áreas amostradas na região do no Pantanal (áreas inundáveis) daquelas que ocorrem em outras localidades do Centro-Oeste (áreas secas). Apesar de que nas análises de Castro (1994), os cerrados do Pantanal e do Centro-Oeste foram agrupados floristicamente no mesmo super-centro de diversidade (Planalto Central), observou-se pelos resultados desse estudo que houve dessemelhança florística entre os cerradões do Pantanal e outros do Centro-Oeste. A inundação pode ter influenciado esta heterogeneidade florística, já que, as áreas analisadas no Distrito Federal, Goiás e aquelas que não fazem parte da planície de inundação em Mato Grosso apresentam situações de precipitação e altitude diferenciadas. A precipitação concentrada no verão em conjunto com as características da declividade do relevo causa o transbordamento dos rios que ocasiona inundações periódicas no Pantanal (Nunes da Cunha & Junk 2009). Este fato provavelmente lhe confere diferenças florísticas em relação às outras áreas do Planalto Central. Junk et al. (1989) discutem que a saturação hídrica limita o estabelecimento de espécies no Pantanal e por isso somente poucas espécies estão adaptadas para esta condição especial que se alterna entre estresse de seca e cheia.
A vegetação estudada sobre cordilheira apresentou maior relação florística e estrutural com os cerradões do que com os cerrados s.s. do Centro-Oeste, podendo, dessa forma, ser caracterizada como uma fitofisionomia de cerradão. E conforme as características apresentadas nessa comunidade ela foi identificada como uma savana florestada, embasado no sistema de classificação de habitats do Pantanal proposto por Nunes da Cunha et al. (2006).
Além disso, os cerrados do Pantanal apresentaram menor relação florística com os de outras regiões do Centro-Oeste, essa heterogeneidade florística pode estar relacionada ao pulso de inundação que atua como filtro para a maioria das espécies das outras áreas do Planalto Central.
A identificação florística e estrutural dos cerradões podem ser consideradas como o passo inicial para incrementar o conhecimento sobre a diversidade florística e justificar medidas de manejo e conservação para essas comunidades. Pois estudos que sublinham os cerradões no Pantanal ainda são insuficientes, visto que este tipo florestal ocorre geralmente sobre as cordilheiras e segundo Nunes da Cunha & Junk (1999), são áreas extremamente visadas para a pecuária e estão sendo destruídas sem que se tenha o conhecimento de seus recursos naturais. Adicionalmente a investigações florística para outras áreas, recomenda-se ainda, estudos que investiguem as relações da vegetação com os fatores abióticos que poderão subsidiar a elaboração de normas específicas para uso destes habitats.
Agradecimentos
Este artigo dedicamos a Profa. Dra. Jeanine Felfili falecida neste ano. Ao Projeto Ecologia do Pantanal, programa SHIFT, cooperação bilateral Brasil e Alemanha e ao Centro de Pesquisas do Pantanal (CPP) pelo financiamento logístico. CNPq pela concessão de bolsa (DTI). A Universidade Federal de Mato Grosso, Instituto de Biociências, Programa de Pós Graduação em Ecologia e Conservação da Biodiversidade pelo apoio científico concedido. Leandro Lima de Sousa pelo abstract. Ao proprietário da Fazenda Nossa Senhora Aparecida, Sr. Carlos Maciel (Sr. Titi) e a toda comunidade do Pirizal.
Recebido em 23/11/2009
Versão reformulada recebida em 14/06/2010
Publicado em 07/07/2010
- AB'SABER, A. N. 1983. Os domínios dos cerrados: introdução ao conhecimento. Rev. Servidor Público 111:41-55.
- ANGIOSPERM PHYLOGENY GROUP. 2009. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG III. Bot. J. of the Linn. Soc. 161:105-121.
- ASSUNCAO, S.L. & FELFILI, J.M. 2004. Fitossociologia de um fragmento de cerrado sensu stricto na APA do Paranoá, DF, Brasil. Acta Bot. Bras. 18:903-909.
- BALDUINO, A.P.C., SOUZA, A.L., MEIRA NETO, A.A., SILVA, A.F. & SILVA Jr., M.C. 2005 Fitossociologia e análise comparativa da composição florística do cerrado da flora de Paraopeba-MG. Rev. Árvore 29:25-34.
- BARBOSA, R.I., NASCIMENTO, S.P., AMORIM, P.A.F. & SILVA, R.F. 2005. Notas sobre a composição arbóreo-arbustiva de uma fisionomia das savanas de Roraima, Amazônia Brasileira. Acta Bot. Bras. 19:323-329.
- BATALHA, M.A. & MARTINS, F.R. 2007. The vascular flora of the cerrado in Emas National Park (Central Brazil): a savanna flora summarized. Braz. Arch. Biol. Technol. 50:269-277.
- BORGES, H.B.N. & SHEPHERD, G.J. 2005. Flora e estrutura do estrato lenhoso numa comunidade de Cerrado em Santo Antônio do Leverger, MT, Brasil. Rev. Brasil. de Bot. 26:61-74.
- CASTRO, A.A.J.F. 1994. Comparação florístico-geográfica (Brasil) e fitossociológica (Piauí - São Paulo) de amostras de cerrado. Tese de Doutorado, Universidade Estadual de Campinas, Campinas, SP, 520p.
- CASTRO, A.A.J.F., MARTINS, F.R., TAMASHIRO, J.Y. & SHEPHERD, G.J. 1999. How rich is the flora of Brazilian cerrados? Ann. Missouri Bot. Gard. 86:192-224.
- DAMASCENO Jr., G.A.; BEZERRA, M.A.O.; BORTOLOTTO, I.M.; POTT, A. 1999. Aspectos florísticos e fitofisionômicos dos capões do Pantanal do Abobral. In: SIMPÓSIO SOBRE RECURSOS NATURAIS E SÓCIO-ECONÔMICOS DO PANTANAL, 2., 1996, Corumbá, MS. Manejo e Conservação. Anais... Corumbá: EMBRAPA Pantanal, p. 203-214.
- DAMASCENO Jr., G.A, SEMIR, J., SANTOS, F.M. & LEITÃO-FILHO, H.F. 2004. Tree mortality in a riparian forest at Rio Paraguai, Pantanal, Brazil, after an extrreme flooding. Acta Bot. Bras. 18:839-846.
- DAMASCENO Jr., G.A., SEMIR, J., SANTOS, F.A.M. & LEITÃO-FILHO, H.F. 2005. Structure, distribution of species and inundation in a riparian forest of Rio Paraguai, Pantanal, Brazil. Flora 200:119-135.
- DUBS, B. 1994. Differentiation of woodland and wet savanna habitats in the Pantanal of Mato Grosso. Brasil. Betrona-Verlag, Küsnacht.
- EITEN, G. 1982. Brasilian "Savannas". In Ecology of Tropical Savannas: Ecological Studies (Huntley, B.J. & Walker, B.H., eds.). Spinger Verlag, Berlin, p. 25-47.
- EITEN, G. 1994. Vegetação do cerrado. In Cerrado: caracterização, ocupação e perspectivas (M.N. Pinto, ed.). EDUNB; SEMATEC, Brasília, DF, p. 17-73.
- FELFILI, J.M. & SILVA Jr., M.C. 1993. A comparative study of cerrado (sensu stricto) vegetation in Central Brazil. J. Trop. Ecol. 9:277-289.
- Felfili, J.M. & Silva Júnior, M.C. 1998. Distribuição dos diâmetros numa faixa de cerrado na Fazenda Água Limpa (FAL) em Brasília - DF. Acta Bot. Brasil. 2:85-104.
- FELFILI, J.M., NOGUEIRA, P.E., SILVA Jr., M.C., MARIMON, B.S. & DELLITTI, W.B. 2002. Composição florística e fitossociologia do cerrado sentido restrito no município de Água Boa - MT. Acta Botanica Brasilica 16(1):103-112.
- FELFILI, J.M., REZENDE, A.V., SILVA Jr., M.C. & SILVA, M.A. 2000. Changes in the floristic composition of cerrado sensu stricto in Brazil over a nine-year period. J. Trop. Ecol. 16:579-590.
- FELFILI, J.M., REZENDE, A.V., SILVA Jr., M.C., SILVA, P.E.N., WALTER, B.M.T., ENCINAS, J.I. & SILVA, M.S. 2007. Fitossociologia da vegetação arbórea. In Biogeografia do bioma Cerrado: vegetação e solos da Chapada dos Veadeiros (J.M. Felfili, A.V. Rezende & M.C. Silva Jr., org.). Universidade de Brasília, Departamento de Engenharia Florestal, Brasília, DF, p. 47-110.
- FELFILI, J.M., SILVA Jr., M.C., REZENDE, A.V., MACHADO, B.W.T., SILVA, P.E.N. & HAY, J.D. 1993. Análise comparativa da florística e fitossociologia da vegetação arbórea do cerrado sensu stricto na Chapada Pratinha, Brasil. Acta Bot. Brasil. 6:27- 46.
- FELFILI, J.M., SILVA Jr., M.C., SEVILHA, A.C., REZENDE, A.V., NOGUEIRA, P.E., WALTER, B. M. T., CHAGAS, E., SILVA, F. & SALGADO, A.S. 2001. Fitossociologia da vegetação arbórea. In Biogeografia do bioma Cerrado: estudo fisionômico da Chapada do Espigão Mestre do São Francisco (J.M. Felfili & M.C. Silva Jr. org.). Universidade de Brasília, Departamento de Engenharia Florestal, Brasília, DF, p. 35-56.
- FISHER, R.A., CORBET, A.S. & WILLIAMS, C.B. 1943. The relationship between the number of species and the number of individuals in a random sample of an animal population. J. Anim. Ecol. 12:42-58.
- FORMAN, R.T.T. & GODRON, M. 1986. Landscape Ecology. John Wiley & Sons, New York, 619p.
- GONZALEZ-RIVAS, B., TIGABU, M., GERHARDT, K., CASTRO-MARIN, G. & ODÉN, PC. 2006. Species composition, diversity and local uses of dry deciduous and gallery forests in Nicaragua. Biod. & Cons. 15:1509-1527.
- GOODLAND, R.J.A. 1970. Plants of the cerrado vegetation of Brasil. Phytologia 20:57-78.
- GOTTSBERGER, G. & SILBERBAUER-GOTTSBERGER, I. 2006. Life in the Cerrado: a South American Tropical Seasonal Vegetation. Vol. I: Origin, Structure, Dynamics and Plant Use. Reta Verlag, Ulm.
- GUARIM NETO, G., GUARIM, V.L.M.S., MORAES, E.C.C., FERREIRA, L.A.D. 1996. Fitossociologia de matas ciliares no Pantanal Mato-Grossense. Bol. Mus. Para. Emilio Goeldi Série Botânica 12:251-263.
- GUARIM, V.L.M.S., MORAES, E.C.C, PRANCE, G.T. & RATTER, J.A. 2002. Inventory of a mesotrophic Callisthene cerradão in the Pantanal of Mato Grosso, Brazil. Edinb. J. Bot. 57:429-436.
- HERINGER, E.P., BARROSO, G.M., RIZZO, J.A. & RIZZINI, C.T. 1977. A flora do cerrado. In IV Simpósio sobre o cerrado: Bases para a utilização agropecuária (M.G. Ferri, org.). Edusp, São Paulo, p. 211-232.
- HILL, M.O. & GAUCH, H.G. 1980. Detrended Correspondence Analysis: An Improved Ordination Technique. Vegetatio 42:47-58.
- ISHARA, K.L., DESTRO, G.F.G., MAIMONI-RODELLA, R.C.S. & YANAGIZAWA, Y.A.N.P. 2008. Composição florística de remanescente de cerrado sensu stricto em Botucatu, SP. Rev. Brasil. Bot. 31:75-586.
- JARDIM, A., KILLEEN, T.J. & FUENTES, A. 2003. Guia de los arboles y arbustos del Bosque Seco Chiquitano, Bolívia. Editorial FAN, Santa Cruz de la Sierra, 324p.
- JUNK, W.J., BAYLEY, P.B. & SPARKS, R.E. 1989. The flood pulse concept in river- foodplain systems. In Proceedings International Large River Symposium (LARS) (D.P. Dodge, ed.). Can. Spec. Pub. Fish. Aquat. Sci. 106:110-127.
- MAGURRAN, A.E. 1988. Ecological Diversity and its Measurement. Cambridge University Press, London, 179p.
- MARIMON Jr., B.H. & HARIDASAN, M. 2005. Comparação da vegetação arbórea e características edáficas de um cerradão e um cerrado sensu stricto em áreas adjacentes sobre solo distrófico no leste de Mato Grosso, Brasil. 2005. Acta Bot. Brasil. 19(4):913-926.
- MARTINS, F.R. 1991. Estrutura de uma floresta mesófila. Ed. UNICAMP, Campinas, 246p.
- McCUNE, B. & MEFFORD, M.J. 1999. PC-ORD version 4.0; Multivariate analysis of ecological data; Users guide. Glaneden Beach: MjM Software Design, 237 p.
- MISSOURI BOTANICAL GARDEN. W3TRÓPICOS. http://www.mobot.org (último acesso em 02/2010.
- MORI, S.A., BOOM, B.M., CARVALINO, A.M. & SANTOS, T.S. 1983. Ecological importance of Myrtaceae in a eastern Brazilian wet forest. Biotropica 15:68-70.
- MÜLLER-DOMBOIS, D. & ELLENBERG, H. 1974. Aims and methods in vegetation ecology. John Wiley and Sons, New York.
- NASCIMENTO, M.T. & SADDI, N. 1992. Structure and floristic composition in an area of cerrado in Cuiabá -MT, Brazil. Rev. Brasil. Bot. 15:47-55.
- NERI, A.V., MEIRA NETO, J.A.A., SILVA, A.F., MARTINS, S.V. & SAPORETTI Jr., A.W. 2007. Composição florísitca de uma área de cerrado stricto sensu no município de Senador Modestino Gonçalves, Vale do Jequitinhonha (MG) e análise de similaridade florística de algumas áreas de cerrado em Minas Gerais. Rev. Árvore 31:1109-1119.
- NUNES DA CUNHA, C. & JUNK, W.J. 1999. Composição florística de capões e cordilheiras: Localização de espécies lenhosas quanto ao gradiente de inundação do Pantanal de Poconé, MT - Brasil. In Anais do II Simpósio sobre recursos naturais e sócio-econômicos do Pantanal - Manejo e Conservação. Corumbá, MS.
- NUNES DA CUNHA, C. & JUNK, W.J. 2001. Distribution of wood plant communities along the flood gradient in the Pantanal of Poconé, Mato Grosso, Brazil. Int. J. Ecol. Environ. Sci. 27:63-70.
- NUNES DA CUNHA, C. & JUNK, W.J. 2004. Year-to-year changes in water level drive the invasion of Vochysia divergens in Pantanal grasslands. Appl. Veg. Sci. 7:103-110.
- NUNES DA CUNHA, C. & JUNK, W.J. 2009. A preliminary classification of habitats of the Pantanal of Mato Grosso and Mato Grosso do Sul, and its relation to national and international wetland classification systems. In The Pantanal: ecology, biodiversity and sustainable management of a large neotropical seasonal wetland (W.J. Junk, C.J. Da Silva, C. Nunes da Cunha & K.M. Wantzen, eds). p. 127-141.
- NUNES DA CUNHA, C., RAWIEL, C.P., WANTZEN, K.M., JUNK, W.J. & LEMES DO PRADO, A. 2006. Mapping and characterization of vegetation units by means of Landsat imagery and management recommendations for the Pantanal of Mato Grosso (Brazil), north of Poconé. Amazoniana 29:1-32.
- OLIVEIRA FILHO, A.T. 1984. Estudo florístico e fitossociológico em um cerrado na Chapada dos Guimarães - Mato Grosso uma análise de gradientes. Dissertação de Mestrado, UNICAMP, Campinas, SP.
- OLIVEIRA-FILHO, A.T. & MARTINS, F.R. 1986. Distribuição, caracterização e composição florística das formações vegetais da região da Salgadeira, na Chapada dos Guimarães (MT). Rev. Brasil. Bot., 9:207-223.
- OLIVEIRA-FILHO, A.T., JARENKOW, J.A. & RODAL, M.J.N. 2006. In Neotropical savannas and seasonally dry forests: plant diversity, biogeography and conservation (R.T. Pennington, G.P. Lewis & J.A. Ratter, eds.). Francis & Taylor, Boca Raton, p. 159-192.
- PAGANO, S.N., LEITÃO FILHO, H.F. & CAVASSAN, O. 1995. Analise Temporal da Composição Florísitca e da Estrutura Fitossociológica de Uma Floresta Mesofila Semidecidua - Rio Claro, Sp. Rev. Brasil. Biol. 55(2):241-258.
- POTT, A. & POTT, V.J. 1994. Plantas do Pantanal. Embrapa- CPAP, Corumbá
- POTT, A. & POTT, V.J. 1999. Flora do Pantanal - Listagem de fanerógamas. In Anais do II Simpósio sobre Recursos Naturais e Sócio-econômicos do Pantanal, Manejo e Conservação. Corumbá, MS, p.297-335.
- PRADO, A.L., HECKMAN, C.W. & MARTINS, F.R. 1994. The seasonal succession of biotic communities in wetlands of the tropical wet-and-dry climatic zone: II. The aquatic macrophyte vegetation in the Pantanal of Mato Grosso, Brazil. Inte. Rev. gesamten Hydrobiol. 79(4):569-589.
- PRANCE, G.T. & SCHALLER, G.B. 1982. Preliminary study of some vegetation types of the Pantanal, Mato Grosso, Brazil. Brittonia 34:228-251.
- RATTER, J.A. 1971. Some notes on two types of cerradão occurring in north eastern Mato Grosso. In III Simpósio sobre o cerrado (M.G. Ferri, coord.). Editora Edgard Blücher Ltda; Editora da Universidade de São Paulo - Edusp, São Paulo, p. 100-103.
- RATTER, J.A., BRIDGEWATER, S. & RIBEIRO, J.F. 2003. Analysis of the floristic composition of the Brazilian cerrado vegetation III: comparison of the woody vegetation of 376 areas. Edinb. J. Bot. 60:57-109.
- RATTER, J.A., BRIDGEWATER, S. & RIBEIRO, J.F. 2006. Biodiversity patterns of woody vegetation of the Brazilian Cerrado. In Neotropical savannas and seasonally dry forests: plant diversity, biogeography and conservation (R.T. Pennington, G.P. Lewis & J.A. Ratter, eds.). Francis & Taylor, Boca Raton, p. 31-66.
- RATTER, J.A., BRIDGEWATER, S., ATKINSON, R. & RIBEIRO, J.F. 1996. Analysis of the floristic composition of the Brazilian cerrado vegetation II: comparison of the woody vegetation of 98 areas. Edinb. J. Bot. 53:153-180.
- RATTER, J.A., POTT, A., POTT, V.J., CUNHA, C.N. & HARIDASAN, M. 1988. Observations on woody vegetation types in the Pantanal and at Corumbá, Brazil. Notes R. Bot. Gard. Edinb. 45:503-525.
- REBELLATO, L. & NUNES DA CUNHA, C. 2005. Efeito do "fluxo sazonal mínimo da inundação" sobre a composição e estrutura de um campo inundável no Pantanal de Poconé, MT, Brasil. Acta Bot. Brasil. 19:789-799.
- RIBEIRO, J.F., SILVA, J.C.S. & BATMANIAN, G.J. 1985. Fitossociologia de tipos fisionômicos de cerrado em Planaltina, DF. Rev. Brasil. Bot. 8:131-142.
- RIBEIRO, L.F. & TABARELLI, M. 2002. A structural gradient in cerrado vegetation of Brazil: changes in woody plant density, species richness, life history and plant composition. J. Trop. Ecol. 18:775-794.
- RIZZINI, C.T. 1997. Tratado de fitogeografia do Brasil: aspectos ecológicos, sociológicos e florísticos. Âmbito Cultural Edições Ltda., Rio de Janeiro.
- SALIS, S.M., ASSIS, M.A., CRISPIM, S.M.A. & CASAGRANDE, J.C. 2006. Distribuição e abundância de espécies arbóreas em cerradões no Pantanal, Estado do Mato Grosso do Sul, Brasil. Rev. Brasil. Bot. 29(3):339-352.
- SALIS, S.M., POTT, V.J. & POTT, A. 1999. Fitosociologia de formações arbóreas da bacia do Alto Paraguai, Brasil. II Simpósio sobre recursos naturais e sócio-econômicos do Pantanal - Manejo e Conservação. Corumbá, MS, 1996.
- SARMIENTO, G. 1984. The ecology of neotropical savannas. Harvard University Press, Cambridge.
- SASAKI, D. & MELLO-SILVA, R. 2008. Levantamento florístico no cerrado de Pedregulho, SP, Brasil. Acta Bot. Bras. 22:187-202.
- SCHESSL, M. 1999. Floristic composition and structure of floodplain vegetation in northern Pantanal of Mato Grosso, Brasil. Phyton 39:303-336.
- SCOLFORO, J.R.S. 1998. Manejo florestal. UFLA/FAEPE, Lavras, 443p.
- SILVA, J.F., FARINÃS, M.R., FELFILI, J.M. & KLINK, C.A. 2006. Spatial heterogeneity, land use and conservation in the cerrado region of Brazil. J. Biogeog. 33:536-548.
- SILVA, M.P., MAURO, R., MOURÃO, G.E. & COUTINHO, M. 2000. Distribuição e quantificação de classes de vegetação do Pantanal através de levantamento aéreo. Rev. Brasil. de Bot. 23:143-152.
- STURGES, H. A. 1926. The choice of a class interval. J. l Am. Stat. 21:65-66.
- TEIXEIRA, M.I.J.G., ARAUJO, A.R.B., VALERI, S.V. & RODRIGUES, R.R. 2004. Florística e fitossociologia de área de cerrado s.s. no município de Patrocínio Paulista, nordeste do Estado de São Paulo. Bragantia 63:1-11.
- TER BRAAK, C.J.F. Ordination. 1995. In: Data analysis in community and landscape ecology (R.H.G. Jongman, C.J.F. Ter Braak & O.F.R. Van Tongeren). Cambridge University Press, Cambrigde, p. 91-173.
- THE PLANT NAMES PROJECT. 2000. International Plant Names Index. http://www.ipni.org (último acesso em 02/2010).
- UHL, C. & MURPHY, P.G. 1981. Composition, Structure and Regeneration of a terra firme Forest in the Amazonian Basin of Venezuela. Trop. Ecol. 22:219-23.
Datas de Publicação
-
Publicação nesta coleção
01 Dez 2010 -
Data do Fascículo
Set 2010
Histórico
-
Recebido
23 Nov 2009 -
Aceito
07 Jul 2010 -
Revisado
14 Jun 2010