Abstracts
This systematic study of the polychaete species of the family Glyceridae is part of a broader program to inventory and determine parameters for conservation and sustainable use of the marine biodiversity off the southeastern and southern coast of Brazil. The material studied was collected from intertidal sandy beaches, the inner continental shelf (<50 m depth) off the northern coast of the state of São Paulo, and the outer continental shelf and part of the continental slope (depths from 60 to 808 m) off the state of Rio de Janeiro south to Rio Grande do Sul (22° S - 34° 40' S). The descriptions of several species were improved, adding new and important taxonomic characters, mainly related to proboscideal papillae. A key for identification of the glycerid species from southeastern and southern Brazil is also provided. Eight species of Glyceridae were identified: Glycera americana Leidy 1855; Glycera brevicirris Grube 1870; Glycera dibranchiata Ehlers 1868; Glycera lapidum Quatrefages 1866; Glycera oxycephala Ehlers 1887; Glycera tesselata Grube 1863; Hemipodia californiensis (Hartman 1938); Hemipodia simplex (Grube 1857); and a previously unknown species of Glycera.
Glyceridae; Glycera; Hemipodia; new species; new occurrence; Brazil
O estudo sistemático das espécies da família Glyceridae é parte de um amplo programa de inventariamento e determinação de parâmetros abióticos para conservação e uso sustentável da biodiversidade marinha ao largo da costa sudeste e sul do Brasil. O material estudado foi coletado desde a região entremarés de praias arenosas até a plataforma interna (<50 m prof.) na costa norte de São Paulo, e da plataforma externa ao talude superior (profundidades entre 60 e 808 m) desde a costa sul do Rio de Janeiro até a o sul do Rio Grande do Sul (22° S - 34° 40' S). As descrições de algumas espécies foram ampliadas, com a adição de novos e importantes caracteres taxonômicos, principalmente relacionados às papilas proboscidiais. Uma chave de identificação para as espécies de glicerídeos da região sudeste e sul do Brasil é fornecida. Oito espécies de Glyceridae foram identificadas: Glycera americana Leidy 1855; Glycera brevicirris Grube 1870; Glycera dibranchiata Ehlers 1868; Glycera lapidum Quatrefages 1866; Glycera oxycephala Ehlers 1887; Glycera tesselata Grube 1863; Hemipodia californiensis (Hartman 1938); Hemipodia simplex (Grube 1857); além de uma espécie previamente desconhecida de Glycera.
Glyceridae; Glycera; Hemipodia; espécie nova; nova ocorrência; Brasil
ARTICLES
Glyceridae Grube 1850 (Annelida: Polychaeta) from Southern and Southeastern Brazil, including a new species of Glycera
Glyceridae Grube 1850 (Annelida: Polychaeta) da região sudeste e sul do Brasil, incluindo uma nova espécie de Glycera
Alexandra Elaine Rizzo11 Corresponding author: Alexandra Elaine Rizzo, e-mail: aerizzo@hotmail.com; Tatiana Menchini Steiner; Antonia Cecília Zacagnini Amaral
Instituto de Biologia IB, Zoologia, Universidade Estadual de Campinas Unicamp, CP 6109, CEP 13083-970, Campinas, SP, Brazil, e-mail: tatims@unicamp.br, http://www.ib.unicamp.br/
ABSTRACT
This systematic study of the polychaete species of the family Glyceridae is part of a broader program to inventory and determine parameters for conservation and sustainable use of the marine biodiversity off the southeastern and southern coast of Brazil. The material studied was collected from intertidal sandy beaches, the inner continental shelf (<50 m depth) off the northern coast of the state of São Paulo, and the outer continental shelf and part of the continental slope (depths from 60 to 808 m) off the state of Rio de Janeiro south to Rio Grande do Sul (22° S - 34° 40' S). The descriptions of several species were improved, adding new and important taxonomic characters, mainly related to proboscideal papillae. A key for identification of the glycerid species from southeastern and southern Brazil is also provided. Eight species of Glyceridae were identified: Glycera americana Leidy 1855; Glycera brevicirris Grube 1870; Glycera dibranchiata Ehlers 1868; Glycera lapidum Quatrefages 1866; Glycera oxycephala Ehlers 1887; Glycera tesselata Grube 1863; Hemipodia californiensis (Hartman 1938); Hemipodia simplex (Grube 1857); and a previously unknown species of Glycera.
Keywords: Glyceridae, Glycera, Hemipodia, new species, new occurrence, Brazil.
RESUMO
O estudo sistemático das espécies da família Glyceridae é parte de um amplo programa de inventariamento e determinação de parâmetros abióticos para conservação e uso sustentável da biodiversidade marinha ao largo da costa sudeste e sul do Brasil. O material estudado foi coletado desde a região entremarés de praias arenosas até a plataforma interna (<50 m prof.) na costa norte de São Paulo, e da plataforma externa ao talude superior (profundidades entre 60 e 808 m) desde a costa sul do Rio de Janeiro até a o sul do Rio Grande do Sul (22° S - 34° 40' S). As descrições de algumas espécies foram ampliadas, com a adição de novos e importantes caracteres taxonômicos, principalmente relacionados às papilas proboscidiais. Uma chave de identificação para as espécies de glicerídeos da região sudeste e sul do Brasil é fornecida. Oito espécies de Glyceridae foram identificadas: Glycera americana Leidy 1855; Glycera brevicirris Grube 1870; Glycera dibranchiata Ehlers 1868; Glycera lapidum Quatrefages 1866; Glycera oxycephala Ehlers 1887; Glycera tesselata Grube 1863; Hemipodia californiensis (Hartman 1938); Hemipodia simplex (Grube 1857); além de uma espécie previamente desconhecida de Glycera.
Palavras-chave: Glyceridae, Glycera, Hemipodia, espécie nova, nova ocorrência, Brasil.
Introduction
The family Glyceridae comprises several species included in a few genera, the largest of which is Glycera Savigny 1818, followed by Hemipodia Kinberg 1865 and Glycerella Arwidsson 1899. Böggemann (2002) revalidated the genus Hemipodia as the senior synonym of Hemipodus Quatrefages 1866. The differences among the three genera are basic: Hemipodia contains species with only uniramous parapodia throughout the body, whereas in Glycera and Glycerella the parapodia are almost all biramous. The ailerons are either rod-like (Glycerella and Hemipodia) or possess a more complicated structure with outer and inner rami, and sometimes an interramal plate (Glycera). Glycera and Hemipodia have spinigerous compound neurochaetae, but in Glycerella additional falcigerous chaetae are present. Böggemann (2002) presented the first revision of all members of Glyceridae previously described worldwide, reducing the total of the 166 recent taxa to only 42 species (36 Glycera, 1 Glycerella and 5 Hemipodia).
Taxonomic studies of several species of Glycera and Hemipodia (=Hemipodus) from the Brazilian coast have been carried out by Nonato & Luna (1970), Orensanz & Gianuca (1974), Rullier & Amoureux (1979), Nonato (1981), and Lana (1984). In addition, Amaral & Nonato (1996) published keys for identification of families and genera of polychaetes from Brazil and Amaral et al. (2006a) a catalogue of polychaete species from Brazil. Until now, 17 species and all three genera are mentioned for the Brazilian coast, but some of them were synonymized by Böggemann (2002).
The aim of this study is to describe and illustrate species of Glyceridae that were collected during projects to develop essential information for the conservation and sustainable use of biodiversity off the Brazilian coast. Eight species of glycerids were identified: Glycera americana Leidy 1855; Glycera brevicirris Grube 1870; Glycera dibranchiata Ehlers 1868; Glycera lapidum Quatrefages 1866; Glycera oxycephala Ehlers 1887; Glycera tesselata Grube 1863; Hemipodia californiensis (Hartman 1938) and Hemipodia simplex (Grube 1857). A new species, Glycera boeggemanni, is described and keys for identification of the species of Glycera and Hemipodia from this region are supplied.
Material and Methods
The examined material are from different programs, sampled in southern and southeastern Brazil (22° S 34°40' S): "Fauna de Praia" (Sandy Beach Fauna), in the intertidal zone of 13 beaches along the São Sebastião Channel (State of São Paulo); "REVIZEE/Score Sul Bentos" (Program of Available of the Sustainable Potential of the Living Resources of the Exclusive Economic Zone - EEZ), in depths from 60 to 808 m, between Ilha Grande bay (Rio de Janeiro State) and Tramandaí city (Rio Grande do Sul State); and "BIOTA/FAPESP - Bentos Marinho" (Benthic Marine Biodiversity in the State of São Paulo), in intertidal sandy beaches and rocky shores and non-consolidated sublittoral (50 m depth), in the northern São Paulo State. The specific methodology to each program can be found, respectively, in Amaral et al. (2003), Amaral et al. (2004) and Amaral et al. (2006b).
The nomenclature used for taxonomic features is based on Böggemann (2002). Semipermanent slides were mounted with a substance containing glycerin as the main component. Measurements and line drawings were made using ZEISS optical microscopy and stereomicroscopy. Scanning electron microscope observations were made at the Laboratório de Microscopia Eletrônica, Instituto de Biologia, Universidade Estadual de Campinas (UNICAMP), with JEOL JSM-5800 LV® equipment, after osmium washing, alcohol battery, critical-point drying with Balzers CPD 30® (37 °C temperature and 70 kg.cm2 pressure) equipment and coating with 44 nm gold. The material examined was deposited at the Museu de História Natural (MHN-UNICAMP) under the abbreviations ZUEC-BPO AR or ZUEC-BPO ST. Others abbreviations used in the material examined: VFS (very fine sand), FS (fine sand), CS (coarse sand).
Additional material from BMNH (The Natural History Museum, London, UK), HZM (Zoologisches Institut und Zoologisches Museum der Universität Hamburg, Germany), SMF (Senckenbergmuseum, Frankfurt am Main, Germany), SSM (Swedish Museum of Natural History, Sweden), ZMB (Museum für Naturkunde der Humboldt Universität zu Berlin, Germany), ZMUC (Zoological Museum, University of Copenhagen, Denmark) was examined as well.
Results and Discussion
Systematics
1. Glycera Savigny 1818
Type species: Glycera unicornis Savigny 1818
Key to species of Glycera from Southeastern-Southern Brazil:
1. Mid-body chaetigers with two prechaetal and one postchaetal lobes; branchiae absent....................................................................................................... 2
- Mid-body chaetigers with two prechaetal and two postchaetal lobes; branchiae present or absent........................................................................................ 3
2. Prechaetal lobes of about same length; ailerons with slightly arched bases................................................................................ Glycera oxycephala
- Notopodial prechaetal lobes distinctly shorter than neuropodial lobes; ailerons with slight dent in pointed triangular bases ........................................ Glycera lapidum
3. Ailerons with deeply incised bases; branchiae absent; digitiform proboscideal papillae (type 1) (Figure 4d).......................................................................... 4
- Ailerons with triangular or rounded triangular bases; branchiae present, conical proboscideal papillae (type 1) (Figure 1b)......................................................... 5
4. Papillae (type 1) with straight, median, longitudinal ridge only..................................................................................... Glycera tesselata
- Papillae (type 1) with 6-20 transverse ridges........................... Glycera brevicirris
5. One retractile branchia per parapodium; ailerons with triangular bases................ 6
- Two branchiae per parapodium (situated dorsally and ventrally on parapodial bases); ailerons with rounded triangular bases.................................... Glycera dibranchiata
6. Papillae (type 1) with 2 ridges; branchiae bush-like................. Glycera americana
- Papillae (type 1) with 5-6 ridges; branchiae with up to six rami............................................................................... Glycera boeggemanni
2. Glycera americana Leidy 1855
(Figure 1)
Glycera americana Leidy 1855:147-148, pl. 11, Figs. 49-50; Nonato & Luna 1970:71-72, Fig. 26; Orensanz & Gianuca 1974:11; Nonato 1981:103-104 (unpublished thesis); Temperini 1981:28-29 (unpublished thesis); Lana 1984: 92-94, Figs. 84-85 (unpublished thesis); Böggemann 2002: 65-66, Figs. 88-90.
Material examined
Type material:Glycera heteropoda Hartmann-Schröder 1962, holotype, HZM P-15330, Chile, Penco, 2.i.1958; Glycera jucunda Kinberg 1865, syntype, SSM Type 6042, Brazil, Rio de Janeiro Harbor, R/V Eugenie 1851-53, 7.28 m; syntype, SSM Type 6043, Brazil, Rio de Janeiro Harbor, R/V Eugenie 1851-53, 7.28 m; Glycera laevis Kinberg 1865, syntype, SSM Type 6030, Brazil, Cabo Frio, 22° 30' S and 40° 55' W, R/V Eugenie 1851-53, 36-55 m; syntype, SSM Type 6031, Brazil, Cabo Frio, 22° 30' and S 40° 55' W, R/V Eugenie 1851-53, 36-55 m; syntype, SSM Type 6040, Brazil, Cabo Frio, 22° 30' S and 40° 55' W, R/V Eugenie 1851-53, 36-55 m; syntype, SSM Type 6041, Brazil, Cabo Frio, 22° 30' S and 40° 55' W, R/V Eugenie 1851-53, 36-55 m.
Additional material: 27 specimens: sta. 6814, 29° 15' 00" S and 48° 41' 80" W, 100 m, 23.III.1999 (ZUECBPOAR15, 1 spec.); sta. 210i, 23° 47' 476" S and 45° 08' 229" W, 20 m, VFS (ZUECBPOAR497, 2); sta. 127i, 23° 31' 837" S and 45° 01' 887" W, 17.3 m, FS (ZUECBPOAR498, 1); sta. 201i, 23° 30' 981" S and 45° 65' 517" W, 8 m, FS (ZUECBPOAR502, 1); sta. 176i, 23° 34' 732" S and 44° 51' 059" W, 20.1 m, CS (ZUECBPOAR504, 2); sta. 163i, 23° 52' 330" S and 45° 34' 227" W, 24.6 m, VFS (ZUECBPOAR505, 1); sta. 161i, 23° 58' 557" S and 45° 31' 013" W, 33.6 m, FS (ZUECBPOAR506, 1); sta. 156i, 23° 46' 955" S and 45° 10' 421" W, 39.3 m, FS (ZUECBPOAR507, 1); sta. 139i, 23° 30' 651" S and 45° 10' 233" W, 6.4 m, VFS (ZUECBPOAR508, 1); sta. 159i, 23° 55' 047" S and 45° 13' 561" W, 27.5 m, VFS (ZUECBPOAR511, 1); sta. 147i, 23° 40' 602" S and 45° 05' 622" W, 35.2 m, VFS (ZUECBPOAR523, 1); sta. 152i, 23° 39' 334" S and 45° 18' 924" W, 15.4 m, VFS (ZUECBPOAR533, 1); sta. 135i, 23° 31' 618" S and 45° 06'358" W, 15.1 m, VFS (ZUECBPOAR537, 1); sta. 94i, 23° 23' 696" S and 44° 52' 113" W, 17.3 m, FS (ZUECBPOAR541, 1); sta. 131i, 23° 33' 745" S and 45° 04' 038" W, 23.4 m, VFS (ZUECBPOAR568, 1); sta. 116i, 23° 26' 193" S and 44° 58' 650" W, 19.8 m, VFS (ZUECBPOAR572, 2); sta. 131i, 23° 33' 745" S and 45° 04' 038" W, 23.4 m, VFS (ZUECBPOAR574, 1); sta. 133i, 23° 32' 539" S and 45° 05' 409" W, 15.4 m, FS (ZUECBPOAR577, 1); sta. 78i, 23° 45' 062" S and 45° 13' 856" W, 25.3 m, FS (ZUECBPOAR585, 1); sta. 141i, 23° 34' 812" S and 45° 09' 386" W, 16.2 m, VFS (ZUECBPOAR586, 1); sta. 50i, 23° 35' 498" S and 45° 17' 879" W, 5.3 m, VFS (ZUECBPOAR588, 1); sta. 46i, 23° 45' 652" S and 45° 03' 309" W, 44 m, FS (ZUECBPOAR590, 1); sta. 8i, 23° 53' 097" S and 45° 30' 845" W, 25 m, VFS (ZUECBPOAR592, 1); sta. 6658, 25° 11' 89" S and 47° 08' 09" W, 157 m, 16.XII.1997 (ZUECBPOAR608, 1).
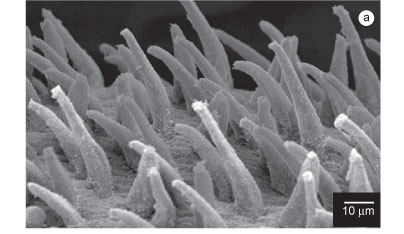
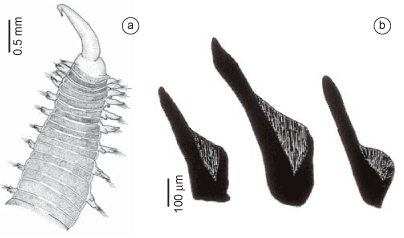
Description (based on additional material): Complete specimens measuring from 110 to 230 mm long, 0.6 to 3.8 mm wide, with 51-257 chaetigers. Coloration yellowish. Brown pigment may be present on prostomial rings, lobes and parapodial cirri. Conical prostomium with 9-12 rings (Figure 1a,f); two specimens (AR 497) with prostomium longer than in others. Nuchal organs dorsolaterally on basal ring. Proboscideal papillae of two types (Figure 1g): type 1. numerous conical papillae anteriorly smooth, posteriorly with two U-shaped ridges (Figure 1b,i) and type 2. isolated, broader papillae without ridges (Figure 1c,j). Aileron with triangular base, interramal plate present (Figure 1d). Branchiae retractile, bush-like when completely developed and everted, starting from about 15th to 26th parapodia and located dorsally on posterior side of parapodial bases (Figure 1e,h). Mid-body segments bi-annulate. First two parapodia uniramous, with a prechaetal and a postchaetal lobe. Following parapodia biramous with two triangular to digitiform prechaetal lobes of about same length, and two shorter postchaetal lobes, anteriorly rounded, posteriorly triangular, of about same length or neuropodial slightly shorter than notopodial (Figure 1e,h). Dorsal cirrus from third parapodium, conical to oval, inserted on body wall slightly above parapodial base. Ventral cirrus from first parapodium, triangular to digitiform, slightly shorter than postchaetal lobes, in posterior parapodia slender to elongated (Figure 1e,h). Last parapodia short and slender. Pygidial cirrus elongated, as long as last ten parapodia. Simple capillary notochaetae and compound spinigerous neurochaetae.
Remarks: The specimens examined by Böggemann (2002) have branchiae located in more anterior chaetigers, between 7 and 22. Glycera cirrata Grube 1857 [partim], Glycera laevis Kinberg 1865, Glycera jucunda Kinberg 1865, Glycera edentata Hansen 1882, and Glycera incerta Hansen 1882 described from specimens collected off Rio de Janeiro were synonymized under Glycera americana by Böggemann (2002). This species has been recorded off the Brazilian coast mainly in ecological studies (Amaral et al. 2006a), and it has been often mistaken for other congeneric species.
Distribution: Atlantic coasts of North and South America, Pacific coasts of South America; Brazil (Alagoas, Sergipe, Rio de Janeiro, São Paulo, Paraná and Rio Grande do Sul). From intertidal region to 120 m depth; in this study, G. americana was collected from 5.3 to 157 m.
3. Glycera boeggemanni new species
Material examined
Type Material: holotype ZUEC BPO-AR581 (23° 44.154' S and 45° 02.007' W, Caraguatatuba, Sta. 21i, 22.IV.2001, 36.1 m); paratype ZUEC BPO-AR496 (23° 53.026' S and 45° 30.386' W, São Sebastião, Sta. 28i, 17.V.2001, 25.6 m).
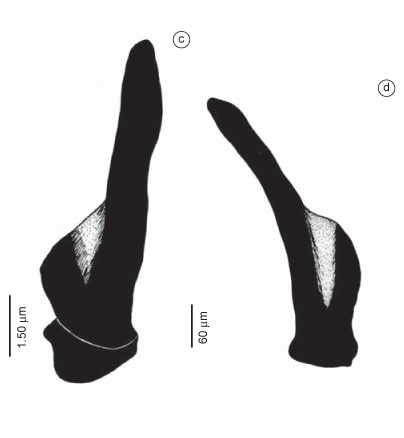
Description: Holotype incomplete, measuring 68 mm long, 3.5 mm wide (invaginated proboscis), with 112 chaetigers. Paratype incomplete, measuring 71 mm long, 3 mm wide (evaginated proboscis), divided into two parts, the anterior with 22-26 chaetigers and the posterior with 68-71 chaetigers. Coloration yellowish to brownish. Pigmentation brownish on dorsum and ventrum, except on corporal rings, lobes and cirri. Conical prostomium with about 9-10 rings, as long as first three chaetigers (Figure 2a). Nuchal organs dorsolaterally on basal ring, drop-shaped. Proboscideal papillae of two types (Figure 2c-d, 3a-d): type 1. numerous conical papillae anteriorly smooth, posteriorly with 5-6 U-shaped ridges (Figure 2c,3a-b, d); type 2. isolated, broader, oval to globular papillae posteriorly with 5-6 ridges in U-shaped (Figure 2d, 3c). Aileron with triangular base (Figure 2b, solid interramal plate indicating a larger specimen). Branchiae retractile, up to six branchial rami, located dorsally at base of parapodia; starting from about parapodium 26 (Figure 2f-h). First segments uni-annulated, following bi-annulated; parapodium and dorsal cirrus inserted on anterior ring. First two parapodia uniramous, with a digitiform prechaetal lobe and short, rounded, postchaetal lobe; dorsal cirrus absent, and ventral cirrus short. Following parapodia biramous, with two digitiform prechaetal lobes of same length, and two short rounded postchaetal lobes of same length (Figure 2e-h). Dorsal cirrus from third parapodium oval, inserted on body wall slightly above parapodial base. Ventral cirrus from first parapodium, triangular, about same length as postchaetal lobes. Parapodia with 7-12 simple capillary notochaetae and 10-16 compound spinigerous neurochaetae, both marginally serrulated (Figures 2 i-j).

Remarks:Glycera boeggemanni n. sp. differs from Glycera robusta Ehlers 1868, its nearest species, mainly in possessing branchiae retractile, up to six branchial rami, in G. boeggemanni n. sp., whereas in G. robusta the branchiae are non-retractile, blister-like; in both species branchiae are located at base on the parapodia. Glycera boeggemanni n. sp. is found in southwest Atlantic, and G. robusta in the western and eastern Pacific Ocean.
Distribution: Brazil (São Paulo: Caraguatatuba and São Sebastião). From 25 to 36 m depth.
Etymology: The species is named after Markus Böggemann, because of his important work and publications on polychaetes, especially glycerids.
4. Glycera brevicirris Grube 1870



Glycera brevicirris Grube 1870:61-62; Böggemann 2002: 44-47, Figs. 34-36
Material examined
Type material:Glycera brevicirris, syntype, ZMB Q. 4304, coll. Grube.
Additional material: 1 specimen: sta.6652, 25° 51' 04" S and 45° 47' 30" W, 206 m, 15.XII.1997 (ZUEC BPO-AR20, preserved for SEM).
Description (based on additional material): Incomplete specimen measuring 13 mm long and 2 mm wide, with 44 chaetigers. Coloration yellowish. Pigmentation brownish, less intense on prostomial ring and more intense on lobes and dorsal cirri of anterior parapodia. Conical prostomium with about 10 rings, as long as first 15 chaetigers (Figure 4a). Drop-shaped nuchal organ on basal prostomial ring. Proboscideal papillae of two types (Figure 4c-d,5a, c-d): type 1. numerous digitiform papillae anteriorly smooth, posteriorly with 8-10 inconspicuous ridges, basally U-shaped and apically V-shaped, and longitudinal lines as intercellular ciliary rootlets; papillae crenulated in lateral view (Figures 4d, 5a, c-d at right); type 2. isolated, broader, smooth -papillae with a straight median longitudinal ridge (Figure 4c, 5d at left). Aileron with deeply incised base (Figure 4b). Branchiae absent. First segments biannulated, with parapodium and dorsal cirrus inserted on posterior ring (Figure 4e, 5b); following segments tri-annulated, with parapodium and dorsal cirrus inserted on middle ring. First two parapodia uniramous, with a prechaetal and postchaetal lobe, dorsal cirrus absent (Figure 5b). Following parapodia biramous, with two triangular to digitiform prechaetal lobes, notopodial slightly shorter than neuropodial lobe (Figure 4e-g, 5b), and two shorter, rounded postchaetal lobes; posterior lobes more elongated and slender (Figure 4g). Dorsal cirrus from third parapodium, digitiform to oval, inserted on body wall slightly above parapodial base (Figure 4e-g,5b). Ventral cirrus from first parapodium, triangular to digitiform, usually longer than the postchaetal lobes and distinctly shorter than the prechaetal lobes (Figure 4e-g). Parapodia with 2-4 capillary simple notochaetae (Figure 4h) and 5-13 spinigerous compound neurochaetae (Figure 4i).
Remarks: The specimens examined by Böggemann (2002) have 10 to 13 prostomial rings and the papillae type 1 has 6 to 20 ridges. Glycera abranchiata, together with Glycera cirrata Grube 1857 [partim], Glycera martensii Grube 1870, ?Glycera fusiformis Fischli 1900, Glycera edwardsi Gravier 1902, Telake epipolasis Chamberlin 1919 and Glycera spadix Treadwell 1943, have been synonymized with G. brevicirris (Böggemann & Fiege 2001; Böggemann 2002). According to Fauchald (1977) and Gilbert (1984), G. abranchiata (=G. brevicirris) was frequently mistaken for G. tesselata, because it was thought that the former possesses two types of proboscideal papillae and the latter only one type. Glycera brevicirris and G. tesselata have both types, basically differing in relation to the digitiform papillae (type 1). In G. brevicirris these papillae have 6-20 ridges, which are basally U-shaped and apically V-shaped, but they may sometimes be inconspicuous (Böggemann 2002:19) or not present when the papillae are poorly preserved or in juvenile specimens, however wrinkles are always present even if the papillae are not curved. In contrast, G. tesselata has digitiform papillae with a straight median longitudinal ridge. The parapodia of G. brevicirris are much more elongated and slender than those of G. tesselata, which are broader and shorter (see description and figures of G. tesselata below). According to Böggemann (2002:23), one syntype of G. cirrata Grube 1857 from Rio de Janeiro, that now is lost, in fact may be G. americana (Augener 1934), while the another syntype from St. Croix (Caribbean Sea) is quite different and should be referred to G. brevicirris.
Distribution: Western and eastern Atlantic, Gulf of Mexico, Caribbean Sea, Red Sea, Indian Ocean, Indo-Pacific, Central Pacific Basin, east Pacific coasts. This study enlarges the distribution of this species, whose southern limit was the Gulf of Mexico, to South America (Brazil, São Paulo). From intertidal zone to 1118 m (Böggemann 2002); in this study, G. brevicirris occurred at 206 m.
5. Glycera dibranchiata Ehlers 1868
Glycera dibranchiata Ehlers 1868:670-702, pl. 24, Figs. 1, 3-8, 10-28; Böggemann 2002:53-54, Figs. 58-60.
Material examined
Additional material: 18 specimens: sta. 2881Pb, 11.XII.1996, 23° 48' 56.06" S and 45° 24' 31.31" W, São Sebastião, Araçá Beach, intertidal (ZUEC-BPO-ST112, 1); sta. 3315Pa, 22.VII.1997, 23° 48' 56.06" , 45° 24' 31.31", São Sebastião, Araçá Beach, intertidal (ZUEC-BPO-ST113, 1); sta. 2749Pa, 27.XI. 1996, 23° 48'56.06", 45° 24' 31.31", São Sebastião, Araçá Beach, intertidal (ZUEC-BPO-ST114, 1); sta. 56M, IX. 1992, 23° 49'04.52" S and 45° 22' 19.95" W, São Sebastião, Perequê Beach, intertidal (ZUEC-BPO-ST110, 1); sta. 95M, III. 1993, 23° 49' 04.52" S and 45° 22' 19.95" W, São Sebastião, Perequê Beach, intertidal (ZUEC-BPO-ST111, 1); sta. 47M, IX. 1992, 23° 49' 09.48" S and 45° 22' 34.88", São Sebastião, Barra Velha Beach, intertidal (ZUEC-BPO-ST109, 1); sta. 149P, 5.IV.2001, 23° 37' 39.4" S and 45° 23' 48.5" W, Caraguatatuba, Camaroeiro Beach, intertidal (ZUEC-BPO-AR108, 1); sta. 234P, 17.IX.2001, 23° 46' 26" S and 45° 40' 30.2" W, São Sebastião, Baleia Beach, intertidal (ZUEC-BPO-AR110, 1); sta. 112P, 4.IV.2001, 23° 37' 30.1" S and 45° 23' 57.1" W, Caraguatatuba, Cidade Beach, intertidal (ZUEC-BPO-AR123, 1); sta. 11P, 13.III.2001, 23° 46' 29.6" S and 45° 41' 39.8" W, São Sebastião, Sahy Beach, intertidal (ZUEC-BPO-AR152, 1); sta. 219i, 23° 49' 955" S and 45° 25' 979" W, 5 m, VFS (ZUEC-BPO-AR500, 1); sta. 153i, 23° 38' 747" S and 45° 20' 397" W, 10 m, VFS (ZUEC-BPO-AR510, 1); sta. 137i, 23° 32' 373" S and 45° 09' 069" W, 10.3 m, VFS (ZUEC-BPO-AR512, 1); sta. 154i, 23° 35' 250" S and 45° 12' 721" W, 5.9 m, VFS (ZUEC-BPO-AR534, 1); sta. 55i, 23° 36' 371" S and 45° 16' 694" W, 8.7 m, VFS (ZUEC-BPO-AR589, 2); sta. 85i, 23° 36' 363" S and 45° 17' 664" W, 12 m, VFS (ZUEC-BPO-AR593, 1); sta. 38i, 23° 23'903" S and 44° 48' 042" W, 24vm, VFS (ZUEC-BPO-AR591, 1).
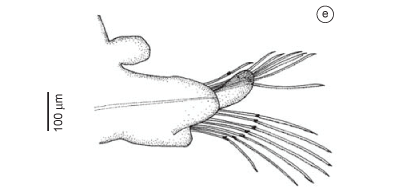
Description: Incomplete specimens measuring from 14 to 140 mm long, 2 to 4.5 mm wide, with 47-182 chaetigers. Coloration from dark yellow to brownish. Conical prostomium with 12-14 rings, as long as the first 10-13 chaetigers (Figure 6a). Nuchal organ a transverse slit and not everted on basal prostomial ring. Proboscis basally with small, oval papillae (Figure 7a), irregularly spaced, with three, almost parallel longitudinal ridge, gradually enlarging towards median and distal region; distally unprovided with papillae (Figure 6b). Proboscideal papillae of two types (Figure 6e-f, 7b-f): type 1, numerous digitiform papillae with 6-7 ridges, U-shaped basally and V-shaped apically (Figure 6e); type 2, isolated, broader, rounded to oval papillae with 3-6 ridges (Figure 6f). Aileron with rounded triangular base; interramal plate small and little translucent (Figure 6c-d). Two simple, digitiform, non-retractile branchiae, starting from about parapodium 16 to 18; upper branchia situated dorsally on parapodial base and lower branchia on anterior side between neuropodial prechaetal lobe and ventral cirrus (Figure 8c-f). Mid-body segments bi-annulate, with parapodium and dorsal cirrus inserted between two rings in anterior region, and on first ring on following segments. First two parapodia uniramous with conical prechaetal lobe, inconspicuous postchaetal lobe and triangular ventral cirrus (Figure 8a). Following parapodia biramous, with two triangular to digitiform prechaetal lobes of about same length (Figure 8b-f), and two shorter postchaetal lobes, more evident from eighth parapodium; shorter, rounded notopodial postchaetal lobe and elongated, triangular neuropodial lobe, both slender in posterior parapodia (Figure 8c-f). Dorsal cirrus from third parapodium, conical to oval, inserted on body wall slightly above parapodial base (Figure 8b). Ventral cirrus from first parapodium, triangular to digitiform, about same length as neuropodial postchaetal lobe (Figure 8b-f). Up to 6 simple capillary notochaetae and 26 compound spinigerous neurochaetae (Figure 8g).

Remarks: The branchiae of specimen ZUEC-BPO-ST111 did not present the fibrous muscular appearance of other specimens; rather, each branchia had a large number of gametes, with the lower branchia almost more inflated (Figure 8f). In some parapodia of this specimen, the branchiae were empty, only the external cuticle remaining. The beginning of the branchiae did not vary much, besides the size differences of the specimens. Böggemann (2002) mentioned 14-17 prostomial rings, branchiae starting from parapodium 13 to 21, and conical proboscideal papillae of type 1 with 4-8 ridges, and of type 2 with 3-6 ridges. A small specimen had the aileron with the outer ramus more curved (Figure 6d).
Distribution: Western and eastern coasts of North and Central America and coast of southern Brazil (São Paulo). From intertidal zone to 403 m; in this study, G. dibranchiata was found from the intertidal zone to 24 m.
6. Glycera lapidum Quatrefages 1866
Glycera lapidum Quatrefages 1866:187-188; Parra et al. 1995:57; Böggemann 2002:37-40, Figs. 19-21.
Material examined
Additional material: 101 specimens: sta. 6762, 23° 26' 20" S and 41° 15' 82" W, 146 m, 28.II.1998 (ZUECBPOAR16, 6); sta. 6644, 25° 45' 80" S and 45° 11' 77" W, 485 m, 14.XII.1997 (ZUECBPOAR21, 1); sta. 6657, 25° 17' 30" S and 46° 55' 60" W, 60 m, 16.XII.1997 (ZUECBPOAR22, 1); sta. 6661, 24° 07' 637" S and 45° 51' 895" W, 147 m, 9.I.1998 (ZUECBPOAR23, 1); sta. 6665, 24° 20' 844" S and 44° 09' 913" W, 258 m, 10.I.1998 (ZUECBPOAR24, 2); sta. 6666, 24° 17' 129" S and 44° 12' 179" W, 163 m, 10.I.1998 (ZUECBPOAR25, 4); sta. 6672, 26° 27' 75" S and 44° 30' 351" W, 165 m, 11.I.1998 (ZUECBPOAR26, 3); sta. 6673, 24° 17' 939" S and 44° 35' 983" W, 133 m, 11.I.1998 (ZUECBPOAR27, 2); sta. 6674, 24° 31' 08" S and 44° 54' 00" W, 122 m, 11.I.1998 (ZUECBPOAR28, 1); sta. 6678, 24° 46' 357" S and 45° 11' 135" W, 99 m, 12.I.1998 (ZUECBPOAR29, 3); sta. 6739, 24° 02' 58" S and 43° 30' 80" W, 147 m, 14.II.1998 (ZUECBPOAR220, 8); sta. 6754, 23° 26' 70" S and 42° 14' 05" W, 131 m, 17.II.1998 (ZUECBPOAR260, 1); sta. 6750, 23° 40' 08" S and 42° 31' 80" W, 162 m, 16.II.1998 (ZUECBPOAR391, 4); sta. 6744, 23° 51' 50" S and 42° 49' 90" W, 254 m, 15.II.1998 (ZUECBPOAR399, 1); sta. 6765, 23° 09' 53" S and 40° 56' 79" W, 257 m, 1.III.1998 (ZUECBPOAR402, 1); sta. 6759, 23° 20' 0" S and 41° 22' 0" W, 110 m, 28.II.1998 (ZUECBPOAR426, 16); sta. 6753, 23° 36' 54" S and 42° 09' 86" W, 187 m, 16.II.1998 (ZUECBPOAR469, 3); sta. 6665, 24° 20' 844" S and 44° 09' 913" W, 258 m, 10.I.1998 (ZUECBPOAR518, 1); sta. 6749, 23° 44' 20" S and 42° 29' 80" W, 325 m, 16.II.1998 (ZUECBPOAR538, 1); sta. 6763, 23° 08' 07" S and 41° 00' 84" W, 101 m, 1.III.1998 (ZUECBPOAR540, 3); sta. 6772, 21° 51' 77" S and 40° 07' 49" W, 110 m, 2.III.1998 (ZUECBPOAR544, 2); sta. 6764, 23° 09' 56" S and 40° 56'19" W, 425 m, 1.III.1998 (ZUECBPOAR546, 1, preserved for SEM); sta. 6765, 23° 09' 53" S and 40° 56' 79" W, 257 m, 1.III.1998 (ZUECBPOAR549, 1); sta. 6769, 22° 02' 87" S and 40° 05' 93" W, 93 m, 2.III.1998 (ZUECBPOAR551, 1); sta. 6762, 23° 26' 20" S and 41° 15' 82" W, 146 m, 28.II.1998 (ZUECBPOAR552, 1); sta. 6750, 23° 40' 08" S and 42° 31' 80" W, 162 m, 16.II.1998 (ZUECBPOAR553, 1); sta. 6753, 23° 36' 54" S and 42° 09' 86" W, 187 m, 16.II.1998 (ZUECBPOAR555, 1); sta. 6761, 23° 29' 04" S and 41° 09' 63" W, 275 m, 28.II.1998 (ZUECBPOAR559, 1); sta. 6747, 23° 17' 30" S and 42° 42' 60" W, 100 m, 16.II.1998 (ZUECBPOAR564, 4); sta. 6741, 23° 49' 90" S and 43° 14' 40" W, 138 m, 15.II.1998 (ZUECBPOAR597, 16); sta. 6673, 24° 17' 939" S and 44° 35' 983" W, 133 m, 11.I.1998 (ZUECBPOAR598, 1); sta. 6761, 23° 29' 04" S and 41° 09' 63" W, 275 m, 28.II.1998 (ZUECBPOAR600, 1); sta. 6739, 24° 02' 58" S and 43° 30' 80" W, 147 m, 14.II.1998 (ZUECBPOAR607, 3); sta. 6657, 25° 17' 30" S and 46° 55' 60" W, 60 m, 16.XII.1997 (ZUECBPOAR609, 1); sta. 6823, 34° 18' 49" S and 52° 02' 76" W, 100 m, 2.IV.1998 (ZUECBPOAR1096, 2); sta. 6844, 32° 38' 03" S and 50° 19' 24" W, 500 m, 5.IV.1998 (ZUECBPOAR1115, 1).
Description: Complete specimens measuring 3.0 to 10.6 mm long (juvenile), 0.3 to 1.2 mm wide, with 25 (juvenile) to 52 chaetigers. Coloration yellowish, without pigments apparent. Conical prostomium, as long as first nine chaetigers, with about 11 rings, inconspicuous and less distinguishable in adults (Figure 9a); juveniles with eight rings. Proboscideal papillae of two types: type 1. numerous, digitiform papillae (Figure 9d, 10a-c), sometimes with inconspicuous undulating ridge; type 2. isolated, shorter and broader, conical to oval papillae without ridges or with an inconspicuous, straight, median longitudinal ridge when completely distended (Figure 9c, 10a-c). Aileron with slight dent in pointed triangular base (Figure 9b). Branchiae absent. Anterior segments (<8) bi-annulated, median and posterior tri-annulated. First two parapodia uniramous; following parapodia biramous, with two triangular to digitiform prechaetal lobes, notopodial lobe usually up to six times smaller than neuropodial lobe; one shorter, rounded postchaetal lobe (Figure 9e-g). Dorsal cirrus from third parapodium, oval to rounded, inserted dorsolaterally on body wall far from parapodial base. Ventral cirrus from first parapodium triangular to digitiform, slightly smaller than postchaetal lobe (Figure 9e-g). Parapodia with 2-3 simple capillary notochaetae (Figure 9h) and 6-11 compound spinigerous neurochaetae (Figure 9i-j). Pygidium rounded; pygidial cirri not observed.
Remarks: Böggemann (2002) mentioned 9-12 prostomial rings for the species. We examined type and non-type material of Glycera capitata Ørsted 1842 [ZMB Q.4306 (?syntypes), SMF 1605, 2173]. Glycera lapidum and G. capitata have only a single postchaetal lobe on all parapodia, the notopodial prechaetal lobe much shorter than the neuropodial lobe, and branchiae absent. The features used to separate these two species are the shape of the proboscideal papillae and ailerons, and the length of the notopodial prechaetal lobe. The shape of the aileron was less variable in the specimens examined here, being nearest to the description of G. lapidum Quatrefages. According to O'Connor (1987), the opening angle of the ramus may vary greatly (45° to 135°). The variations found by this author in the opening angle of both rami at the base, and also in relation to the length of the prechaetal lobe, led him to designate G. lapidum as a complex. However, in the specimens examined here, the opening angle was maintained at about 45° (Figure 9b). In juveniles, because the mandibular apparatus was incompletely developed, the confirmation of the species was based on other characters. The proboscideal papillae observed in our specimens are very similar to those described and illustrated by Böggemann (2002:93, Figure 20), although the undulating ridge of the digitiform papilla (type 1) was not as evident as in his specimens. Observations by SEM and light microscope showed that these ridges may be inconspicuous or even absent in the species of Glycera examined, such as G. americana and G. brevicirris, and this may be indicate that the papillae are poorly preserved or the specimens are juvenile and the papillae are still developing. O'Connor (1987) examined 362 specimens and observed proboscideal papillae of two types, tall with crenate edges and oval; and did not mention any undulating ridge. The length of the prechaetal lobes may be, at first glance, a strong feature to distinguish adult specimens of G. lapidum and G. capitata. In the specimens examined here, the notopodial prechaetal lobe is usually up to six times smaller than the neuropodial prechaetal lobe, corresponding to G. lapidum variety A of O'Connor (1987). This same author suggested that Glycera papillosa Grube 1857 and Glycera mimica Hartman, 1965 may be indistinct from G. lapidum, and that the latter could be a variety of G. lapidum in deep waters. According to Böggemann (2002), G. papillosa, the type material of which is probably lost, is a doubtful name, and may be a junior synonym of G. capitata or G. lapidum; and G. mimica is referred to G. capitata. Glycera kerguelensis McIntosh 1885 and G. mimica were synonymized with G. capitata by Böggemann & Fiege (2001), while G. papillosa was designated as incertae sedis.
Distribution: Mainly in temperate zones and sometimes in tropical seas. This is the first record of this species in Brazil (from São Paulo to Rio Grande do Sul); it had been previously recorded in the South Atlantic (Argentina). From intertidal to 3947 m depth; in this study, G. lapidum occurred from 60 to 500 m.
7. Glycera oxycephala Ehlers 1887
Glycera oxycephala Ehlers 1887:121-123, pl. 41, Figs. 7-11; Parra et al. 1995: 53-59, Figs 1-3; Böggemann 2002: 40-41, Figs. 22-24.
Glycera cf. oxycephala Lana 1984: 94-95, Figs. 86-87.
Material examined: 18 specimens: sta. 6658, 25° 11' 89" S and 47° 08' 09" W, 157 m, 16.XII.1997 (ZUEC-BPO-AR17, 11); sta. 6672, 26° 27' 75" S and 44° 30' 351" W, 165 m, 11.I.1998 (ZUEC-BPO-AR18, 1); sta. 123i, 23° 29' 101" S and 44° 59' 171" W, 24.9 m, AMF (ZUEC-BPO-AR515, 1); sta. 209i, 23° 46' 731" S and 45° 13' 790" W, 12 m, AF (ZUEC-BPO-AR521, 1); sta. 206i, 23° 34' 936" S and 45° 16' 764" W, 3 m, AM (ZUEC-BPO-AR525, 1); sta. 167i, 23° 31' 220" S and 44° 54' 718" W, 44.5 m, AM (ZUEC-BPO-AR528, 1); sta. 45i, 23° 22' 420" S and 44° 53' 135" W, 12 m, AF (ZUEC-BPO-AR591, 1); no number, Paranaguá Bay, Sta. 6110B (1).
Additional material: South Africa, Lambert's Bay St. LAM.26, 32° 04.90' S, 18° 17.50' E, 18.I.1957, 27 m, sand with shells and rocks (BMNH 1963.1.79, 1); Kuwait, Salimiyah and Mena Abdullah, intertidal, sand (BMNH 1971.45, 1)
Description (based on material examined): Incomplete specimens measuring 6 to 23 mm long, 0.6 to 1.8 mm wide, with 35 to 48 chaetigers; only complete specimen (juvenile) with 38 chaetigers. Coloration yellowish, with brownish pigments on dorsum, as transverse bands. Conical prostomium as long as first 12 chaetigers; indistinct rings (Figure 11a). Proboscideal papillae of two types: type 1. numerous conical papillae posteriorly with about 12-13 ridges (Figures 11d,12a-b); type 2. isolated, broader, oval to globular papillae posteriorly with about 8 ridges (Figures 11c,12b); both smooth anteriorly. Aileron with slightly arched base; interramal plate present (Figure 11b). Branchiae absent. Anterior segments bi-annulated, posterior tri-annulated, with parapodium inserted on second ring. Parapodia short, as long as one-tenth of mid-body width. Up to 9 first parapodia uniramous (Figure 11e); following parapodia biramous, with two slender triangular to digitiform prechaetal lobes of about same length, or notopodial lobe slightly longer than neuropodial lobe (Figure 11f-h), and one shorter, rounded postchaetal lobe; both lobes slightly more slender in posterior parapodia. Dorsal cirrus from third parapodium, oval to rounded, inserted on body wall near parapodial base (Figure 11e-h). Ventral cirrus from first parapodium, triangular to digitiform, about as long as postchaetal lobe. Parapodia with 2-5 simple capillary notochaetae and 5-10 compound spinigerous neurochaetae. Pygidial cirri, seen in juvenile, elongated and smooth (Figure 11i).
Remarks: Böggemann (2002) mentioned 20-28 prostomial rings, proboscideal papillae of type 1 with 5-20 ridges, and of type 2 with 4-11 ridges. This species belongs to the group of Glycera with two prechaetal lobes and one postchaetal lobe, and no branchiae. Glycera tenuis Hartman 1944, Glycera ehlersi Arwidsson 1899 and, probably, G. capitata var. benguellana Augener 1931 were synonymized with G. oxycephala by Böggemann & Fiege (2001). The features observed in our specimens agree with the original description of Ehlers (1887), mainly in relation to the prostomium size, reaching the first 12 chaetigers; parapodia 1/12 of mid-body width, becoming more slender and narrow toward last chaetigers; ante-mid-body prechaetal lobes rounded and unequal, except in posterior region, when neuropodial lobe becomes slender and two times longer than notopodial lobe; reduced number of chaetae, about 10, between simple and compound; and position of the dorsal cirrus near parapodial base and not far from parapodial base. However, some features observed here differ from G. tenuis (= G. oxycephala) of Hartman (1944), with about 9 parapodia uniramous, instead of almost all of them, indicating juvenile stages. In juveniles, sometimes all parapodia are uniramous, according to Böggemann (2002: 41). In the specimens examined here, the aileron has a hardened interramal plate. In G. cf. oxycephala of Lana (1984), the aileron may be still in development. The number of ridges varies greatly. Hartman (1968) mentioned proboscideal papillae of one type only, with 13-16 ridges in G. tenuis (= G. oxycephala) and 9-10 ridges in G. oxycephala; Fauchald (1977) found 9-10 ridges and Parra et al. (1995), 8-11. Böggemann & Fiege (2001) and Böggemann (2002) described conical papillae with about 5-20 ridges in G. oxycephala. We found, in our specimens, 12-13 and 8 ridges on the papillae of types 1 and 2, respectively. Differences found on the aileron and proboscideal papillae with 15 rings led to the uncertainty of Lana (1984) as to the assignment to G. oxycephala of his specimens from Paraná state.
Distribution: In temperate zones and tropical seas; Brazil (São Paulo and Paraná). From intertidal zone to 2951 m depth; in this study, G. oxycephala occurred between 3 to 165 m.
8. Glycera tesselata Grube 1863
Glycera tesselata Grube 1863: 41-42, pl. 4, Figs 4, 4a; Böggemann 2002:47-48, Figs 37-39.
Material examined
Type material:Glycera tesselata, syntype, ZMB Q. 4339, Neresine, Lussin piccolo, Croatia, Grube coll.
Additional material: 17 specimens: sta. 6763, 23° 08' 07" S, 41° 00' 84" W, 101 m, 1.III.1998 (ZUEC-BPO-AR563, 1); ZMB Q. 4340 (Italy, Lesina, 15); ZMB 6560 (Bismarck Archipelago, Ralun, 24.XI.1896, 1).
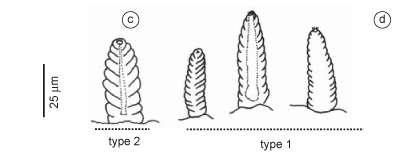
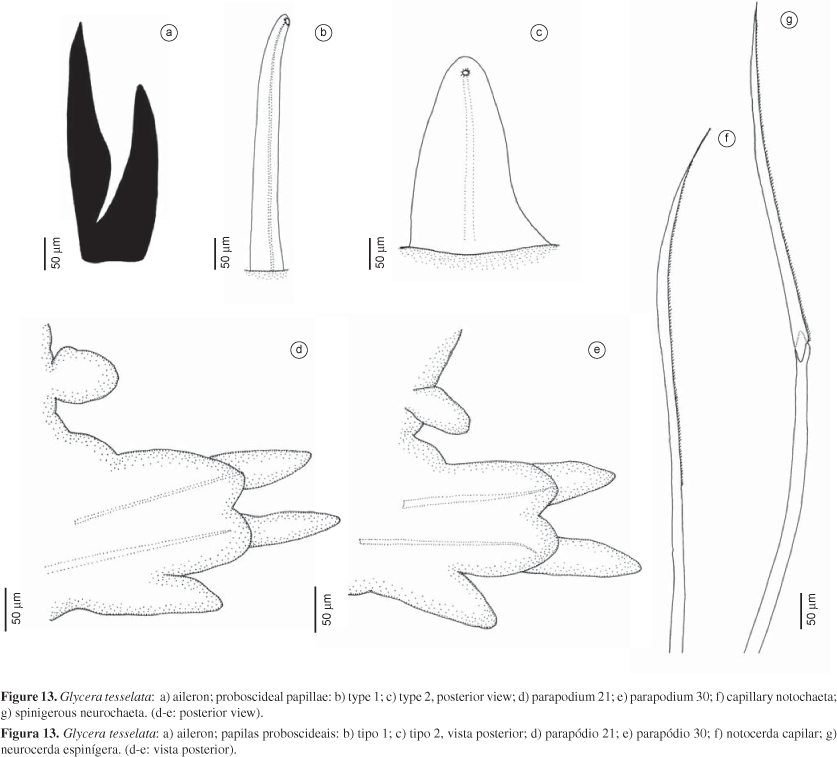
Description (based on additional material AR563): Specimen almost complete, measuring 15 mm long, 1.5 mm wide, with 44 chaetigers. Coloration yellowish; pigment, if present, brownish on prechaetal lobes and ventral cirri. Conical prostomium as long as first 12 chaetigers; indistinct rings. Proboscideal papillae of two types: type 1. numerous digitiform papillae posteriorly with straight, median, longitudinal ridge (Figure 13b); type 2. isolated, shorter and broader, conical papillae, posteriorly with more or less distinctly straight, median, longitudinal ridge (Figure 13c); both papillae smooth anteriorly. Aileron with deeply incised base (Figure 13a). Branchiae absent. Segments bi-annulated, with parapodia inserted on first ring. Parapodia 1/4 mid-body width. First two parapodia uniramous, with single prechaetal and postchaetal lobe; following parapodia biramous, with two triangular to digitiform prechaetal lobes of about same length or notopodial lobe slightly shorter than the neuropodial one on anterior to median parapodia (Figure 13d), or notopodial lobe usually slightly shorter than neuropodial lobe on median to posterior parapodia (Figure 13e); two shorter, rounded postchaetal lobes, about same length; both lobes becoming slender on posterior parapodia. Dorsal cirrus from third parapodium, conical to oval, inserted on body wall not far distant from parapodial base (Figure 13d-e). Ventral cirrus triangular to digitiform, shorter than the postchaetal lobes (Figure 13d-e). Parapodia with simple capillary notochaetae (Figure 13f) and compound spinigerous neurochaeta (Figure 13g).
Remarks: Böggemann (2002) mentioned 8-9 prostomial rings, branchiae absent, and digitiform proboscideal papillae with straight, median, longitudinal ridge. It differs from G. brevicirris, the other congeneric species found in Brazil, in the digitiform proboscideal papillae; G. tesselata has only one straight, median, longitudinal ridge, whereas G. brevicirris has about 6-20 transverse ridges. Our specimen has a dorsal cirrus near the parapodial base, differing from that mentioned by Böggemann (2002:48) for this species, i.e., inserted on the body wall far from the parapodial base mainly on anterior part of the body.
Distribution: Northwestern and northeastern Atlantic, Gulf of Mexico, Caribbean Sea, Mediterranean Sea, Red Sea, south coasts of Africa, Indo-Pacific, northwestern Pacific. This is the first occurrence of the species in Brazil (Rio de Janeiro). From 2 to 4066 m depth; in this study, G. tesselata occurred at 101 m.
9. Hemipodia Kinberg 1865
Type species: Hemipodia simplex (Grube 1857)
Key to species of Hemipodia from southeastern-southern Brazil:
1. Proboscideal papillae of type 1 digitiform with up to 9-40 ridges; all dorsal and ventral cirri rounded to oval ............................................ Hemipodia californiensis
- Proboscideal papillae of type 1 conical only with straight, median, longitudinal ridge; oval ventral cirri on anterior and median chaetigers, elongated and conical cirri on posterior chaetigers ......................................................... Hemipodia simplex
10. Hemipodia californiensis (Hartman 1938)
Hemipodus californiensis Hartman 1938: 93-94, Figs 2-7.
Hemipodus olivieri Orensanz & Gianuca 1974:11-13, Figs 5, 8 and 9; Lana 1984:91, Figs 82-83.
Hemipodia californiensis Böggemann 2002:82, Figs 133 135
Material examined
Additional material: 279 specimens (all samples collected intertidally): sta. 649a/2749Pa/1917a/ 411a/419b/831b/ 833a/645a/651a, 1995 and 1996, 23° 43' S and 45° 25' W, São Sebastião, Enseada Beach, (ZUECBPOST122, ST123, ST124, ST132, ST133, ST134, 12); sta. 2722a/2723a/428b /1825a/1749Pa, 1995 and 1996, 23° 43' 46.25" S and 45° 23' 58.97" W, São Sebastião, Cigarras Beach, (ZUECBPOST138, ST139, ST140, ST141, 5); sta. 682a, 06.XI.1995, 23° 44' 54.29" S and 45° 24' 34.78" W and São Sebastião, São Francisco Beach, (ZUECBPOST136, 1); sta. 2749Pa, 27.XI.1996, 23° 48' 56.06" S and 45° 24' 31.31" W, São Sebastião, Araçá Beach, (ZUECBPOST126, 3); sta. 1777a/694a/698a/ 1048a/1049a/1056a/ 1407a/1601a/1604a/ 1605a/1972a/2155a, 1996, 23° 46' 40.80" S and 45° 23' 59.43" W, São Sebastião, Pontal da Cruz Beach, (ZUECBPOST121, ST125, ST127, ST128, ST129, ST130, ST131, 17); sta. 64a, 07.VIII.1995, 23° 47' 51.94" S and 45° 23' 57.62" W, São Sebastião, Porto Grande Beach, (ZUECBPOST137, 1); sta. 2185a, 03.VII.1996, 23° 49' 22.24" S and 45° 24' 58.42" W, São Sebastião, Grande Beach, (ZUECBPOST135, 1); sta. 110a, X.1995, 23° 49' 40.52" S and 45° 25' 22.61" W, São Sebastião, Segredo Beach, (ZUECBPOST120, 1); sta. 4P/9P/15P/ 11P/10P/14P, 13.III.2001, 23° 46' S and 45° 41' W, São Sebastião, Sahy Beach, (ZUECBPOAR164, AR155, AR153, AR157, AR161, AR167, 15); sta. 16P/20P, 13.III.2001, 23° 46' 27.8" S and 45° 40' 31" W, São Sebastião, Baleia Beach, (ZUECBPOAR151, AR147, 8); sta. 21P/22P/28P, 13.III.2001, 23° 46' 26.6" S and 45° 41' 30.9" W, São Sebastião, Baleia Beach, (ZUECBPOAR131, AR137, AR148, 4); sta. 368P/ 369P/375P, 13.XI.2001, 23° 21' 31.4" S and 44° 51' 23.2" W, Ubatuba, Fazenda Beach, (ZUECBPOAR98, AR96, AR225, 4); sta. 33P/35P, 14.III.2001, 23° 41' 37.8" S and 45° 25' 46.2" W, Caraguatatuba, Palmeiras Beach, (ZUECBPOAR134, AR128, 5); sta. 49P/52P/59P, 14.III.2001, 23° 38' 36.8" S and 45° 25' 23.4" W, Caraguatatuba, Frecheiras Beach, (ZUECBPOAR132, AR163, AR149, AR127, 4); sta. 100P, 3.IV.2001, 23° 37' 21.6", 45° 24' 02", Caraguatatuba, Cidade Beach, (ZUECBPOAR194, 1); sta. 110P/113P/ 116P/117P/ 130P/133P, 4.IV.2001, 23° 37', 45° 23', Caraguatatuba, Cidade Beach, (ZUECBPO AR83, AR150, AR 224, AR 142, AR154, 7); sta. 142P/143P/ 144P/147P/ 148P/149P/ 150P, 5.IV.2001, 23° 37' 39.4", 45° 23' 48.5", Caraguatatuba, Camaroeiro Beach, (ZUECBPOAR158, AR118, AR121, AR169, AR 168, AR 139, AR191, AR185, 10); sta. 151P/152P/ 154P/155P /156P/158P/ 162P, 9.V.2001, 23° 21' 31.1", 45° 51' 21.6", Ubatuba, Fazenda Beach, (ZUECBPOAR82, AR119, AR107, AR171, AR129, AR113, AR214, 7); sta. 169P/173P/ 174P/176P/ 177P/179P, 9.V.2001, 23° 22' 01", 44° 50' 19.7", Ubatuba, Fazenda Beach, (ZUECBPOAR143, AR187, AR212, AR207, AR202, AR192, AR91, 12); sta. 181P/182P/ 183P/184P/ 185P/186P/ 188P, 10.V.2001, 23° 50' 03.8", 45° 30' 38.4", São São Sebastião, ToqueToque Beach, (ZUECBPOAR193, AR100, AR86, AR95, AR126, AR189, AR130, 10); sta. 192P/194P/ 197P/201P/ 202P/204P/ 205P, 21.VIII.2001, 23° 22' S and 44° 50' W, Ubatuba, Picinguaba Beach, (ZUECBPOAR182, AR173, AR186, AR188, AR198, AR197, AR140, AR172, AR174, 16); sta. 208P/210P/ 215P/220P, 17.IX.2001, 23° 46' 29" S and 45° 41' 39.2" W, São Sebastião, Sahy Beach, (ZUECBPOAR99, AR190, AR92, AR177, AR245, 5); sta. 221P/222P/ 224P/226P/ 230P/232P/ 233P/234P, 17.IX.2001, 23° 46' 26" S and 45° 40' 30.2" W, São Sebastião, Baleia Beach, (ZUECBPOAR211, AR205, AR231, AR213, AR248, AR179, AR141, AR124, AR109, AR111, 23); sta. 238P/240P/ 241P/242P/ 243P/244P/ 245P, 19.X.2001, 23° 50' 03.8" S and 45° 30' 38.4" W São São Sebastião, ToqueToque Beach, (ZUECBPOAR84, AR81, AR112, AR115, AR114, AR117, AR209, AR269, AR204, 39); sta. 285P/287P, 19.IX.2001, 23° 38' 15" S and 45° 25' 11.4" W, Caraguatatuba, Frecheiras Beach, (ZUECBPOAR120, AR199, 2); sta. 251P/252P /253P/254P/ 255P/256P/ 257P/258P, 18.IX.2001, 23° 41' 37.4" S and 45° 25' 43.2 W", Caraguatatuba, Palmeiras Beach, (ZUECBPOAR104, AR223, AR106, AR122, AR254, AR165, AR116, AR103, AR101, AR90, 23); sta. 300P/305P, 19.IX.2001, 23° 37' 23.9" S and 45° 24' 21.1" W, Caraguatatuba, Cidade Beach, (ZUECBPOAR183, AR200, 2); sta. 308P/312P/ 324P/325P/ 327P/328P/ 334P, 17.X.2001, 23° 37' S and 45° 24' W, Caraguatatuba, Cidade Beach, (ZUECBPOAR203, AR180, AR305, AR488, AR181, AR201, AR178, AR208 8); sta. 339P/340P/345P/350P, 18.X.2001, 23° 37' 31.9" S and 45° 23' 53.8" W, Caraguatatuba, Cidade Beach, (ZUECBPOAR89, AR94, AR105, AR133, 7); sta. 352P/353P/ 354P/356P/ 357P/358P/ 363P/365P, 18.X.2001, 23° 37' 39.4" S and 45° 23' 48.5" W, Caraguatatuba, Camaroeiro Beach, (ZUECBPOAR184, AR125, AR87, AR88, AR175, AR206, AR195, AR93, 11); sta. 389P/408P, 12 and 13.XI.2001, 23° 22' S and 44° 50' W, Ubatuba, Fazenda Beach, (ZUECBPOAR97, AR236, 2); sta. 67P/69P/70P/73P, 3.IV.2001, 23° 38'15.3" S and 45° 25' 11.2" W, Caraguatatuba, Frecheiras Beach, (ZUECBPOAR146, AR159, AR136, AR176, AR102, 8); sta. 79P/88P/ 90P/91P/ 92P/94P/ 95P/98P, 3.IV.2001, 23° 37' S and 45° 24' W, Caraguatatuba, Cidade Beach, (ZUECBPOAR162/AR170, AR166, AR156, AR145, AR160, AR210, AR135, AR144, 14).
Description: Complete specimens from 12 to 53 mm long, 0.6 to 2.6 mm wide, 83 to 140 chaetigers. Coloration from yellowish to brownish. Conical prostomium with 12 weakly delimited rings, as long as first eight chaetigers. Proboscideal papillae of two types: type 1. numerous digitiform papillae with about 20 U-shaped ridges (Figure 14b-e); type 2. isolated, shorter, slightly broader, more oval to globular papillae with about 15 U-shaped ridges (Figure 14c, e). Aileron rod-like (Figure 14a). Branchiae absent. Anterior segments bi-annulated, with parapodium on first ring, median to posterior tri-annulated, with parapodium on second ring. All parapodia uniramous, with conical prechaetal lobe (Figure 14f), longer than postchaetal lobe, becoming slightly more slender towards posterior end (Figure 14g); postchaetal lobes triangular on anterior parapodia, shorter and blunt on posterior parapodia. Dorsal cirri rounded to oval, inserted on body wall near parapodial base in anterior chaetigers (Figure 14f), becoming far from parapodial base towards posterior (Figure 14g). All ventral cirri rounded to oval, shorter than postchaetal lobe. Up to 16 compound spinigerous chaetae, marginally serrulated (Figure 14h-k).
Remarks: Böggemann (2002) mentioned 8-10 prostomial rings, proboscideal papillae of type 1 with 9-40 ridges, and of type 2 with 7-15 ridges. According to Böggemann (2002), Hemipodus olivieri Orensanz & Gianuca 1974 is synonymous with Hemipodia californiensis. This species is similar to Hemipodia pustatula (Friedrich 1956) in relation to the proboscideal papillae and anterior parapodia. However, H. pustatula has the prechaetal lobe of the posterior chaetiger much more slender and thinner, and the ventral cirrus slender, triangular to digitiform and longer than the postchaetal lobe. Besides this, H. pustatula has a small digitate distal process on the prechaetal lobes starting from the mid-body; such a process is absent in H. californiensis. The smallest specimen examined here is in agreement with the description of juveniles of H. olivieri (= H. californiensis). The largest specimens described by these authors (115 mm long) correspond to the adults described here, but the prechaetal lobes on posterior chaetigers are slightly longer in our specimens. This difference may be due to specimen sizes, because the smaller the specimen, the more elongated is the prechaetal lobe. The same appears to be true for the rings of the proboscideal papillae; i.e., the larger the specimen, the higher the number of rings. Several specimens from 35 mm long had gametes inside the coelomic cavity.
Distribution: West coast of America, from California to Peru, southeastern coasts of South America. From intertidal zone to 100 m depth; common in the intertidal zone of fine-sand beaches in São Paulo state.
11. Hemipodia simplex (Grube 1857)
Glycera simplex Grube 1857: 177-178
Hemipodus rotundus Nonato 1981: 104-106
Hemipodia simplex Böggemann 2002: 79-81, Figs 127-129.
Material examined
Type material: holotype, ZMUC POL-393, Valparaíso, Callao, 21.IX.1841.
Additional material: 13 specimens: sta. 688b, 07.IX.1995, 23° 46' 40.80" S and 45° 23' 59.43" W, São Sebastião, Pontal da Cruz Beach, intertidal (ZUECBPOST119, 1); sta. 63a, 07.VIII.1995, 23° 47' 51.94" S and 45° 23' 57.62" W, São Sebastião, Porto Grande Beach, intertidal (ZUECBPOST117, 1); sta. 62b, 07.VIII.1995, 23° 47' 51.94" S and 45° 23' 57.62" W, São Sebastião, Porto Grande Beach, intertidal (ZUECBPOST118, 2); sta. 200i, 23° 30' 564" S and 45° 08' 340" W, 5.4 m, VFS (ZUECBPOAR509, 5); sta. 116i, 23° 26' 193" Sand 44° 58' 650" W, 19.8 m, VFS (ZUECBPOAR520, 1); sta. 193i, 23° 31' 542" S, 45° 06' 573" W, 9 m, CS(ZUECBPOAR535, 1); sta. 91i, 23° 24' 366" S and 45° 51' 431" W, 16.6 m, AM (ZUECBPOAR557, 1); sta. 92i, 23° 24' 010" S and 44° 50' 902" W, 21.6 m, AM (ZUECBPOAR571, 1).
Description (based on additional material): Complete specimens from 19 to 45 mm long, 1 to 2.2 mm wide, with 88 to 138 chaetigers. Coloration yellowish. Conical prostomium with 10-13 weakly delimited rings, as long as the first nine chaetigers (Figure 15a). Proboscideal papillae of two types (Figure 15c-f): type 1. numerous conical papillae with more or less distinctly straight, median, longitudinal ridge (Figure 15c, e-f); type 2. isolated, broader, oval to globular papillae, without ridges (Figure 15d-f). Aileron rod-like (Figure 15b). Branchiae absent. All parapodia uniramous, with one slender triangular to digitiform prechaetal lobe, distally rounded in anterior chaetigers (Figure 16a-c), becoming longer and thinner towards posterior end (Figure 16d); one shorter, rounded postchaetal lobe. Dorsal cirrus oval and the anteriormost inserted on body wall slightly above parapodial base (Figure 16a-c); slightly shorter and rounded in posterior parapodia (Figure 16d). Ventral cirrus from first parapodium oval, shorter than postchaetal lobe in anterior parapodia (Figure 16a-b), becoming elongated and conical from about chaetiger 70 (Figure 16c), and longer than postchaetal lobe at about chaetiger 120 (Figure 16d). Up to 9 compound spinigerous chaetae, marginally serrated; supra-acicular chaetae with homogomph articulation (Figure 6,h) and subacicular slightly asymmetrical up to parapodium 45 (Figure 16i,j) and symmetrical on following (Figure 16g). In posterior parapodia, blades slightly shorter and slender in relation to the shafts (Figure 16e-g).
Remarks: Böggemann (2002) mentioned 7-9 prostomial rings for the species. According to Grube (1857), the type locality of this species is Valparaiso (Chile) and Callao (Peru). The original description is not very detailed, however, reexamination of the holotype revealed its characteristics in detail. Böggemann (2002) compared specimens identified as Hemipodus rotundus by Nonato (1981) from Ilha Grande Bay (Rio de Janeiro) and verified that it is similar to Hemipodia simplex, as our specimens are also.
Distribution: Cold and warm temperate zone, west and east coasts of North and South America, Bay of Bengal, east coast of Australia, seas around New Zealand. From intertidal zone to 137 m depth; in Brazil, H. simplex occurred from the intertidal zone to 22 m.
Acknowledgments
Our thanks to all participants of the "REVIZEE/South Score", "BIOTA/FAPESP - Marine Benthos" and "Sandy Beach Fauna" Programmes, UNICAMP (Universidade Estadual de Campinas, Instituto de Biologia, Departamento de Zoologia), and at the CEBIMAR/USP (Centro de Biologia Marinha da Universidade de São Paulo). This work was supported by the FAPESP (Fundação de Amparo à Pesquisa, 98/07090-3, 02/04104-0) within the BIOTA/FAPESP - The Biodiversity Virtual Institute Program (www.biotasp.org.br), by the MMA (Ministério do Meio Ambiente, dos Recursos Hídricos e da Amazônia Legal), by the FAEPEX/UNICAMP (Fundo de Apoio ao Ensino, à Pesquisa e à Extensão), and by CNPq (Conselho Nacional de Desenvolvimento Científico e Tecnológico, 380379/02-3, 308072/2006-5). Janet W. Reid revised the English text. We wish to thank also to Emma Sherlock, BMNH (The Natural History Museum, London, UK); Angelika Brandt and Gisela Wegener, HZM (Zoologisches Institut und Zoologisches Museum der Universität Hamburg, Germany); Dieter Fiege, SMF (Senckenbergmuseum, Frankfurt am Main, Germany); Karin Sindemark, SSM (Swedish Museum of Natural History, Sweden); Birger Neuhaus, ZMB (Museum für Naturkunde der Humboldt-Universität zu Berlin, Germany); and Danny Eibye-Jacobsen, ZMUC (Zoological Museum, University of Copenhagen, Denmark) for the loan of specimens.
Data Received 07/02/07
Revised 05/06/07
Accepted 01/09/07
ISSN 1676-0603.
- AMARAL, A.C.Z., DENADAI, M.R., TURRA, A. & RIZZO, A.E. 2003. Intertidal macrofauna in brazilian subtropical tide-dominated sandy beaches. J. Coast. Res. 35:446-455.
- AMARAL, A.C.Z., LANA, P.C., RIZZO, A.E., STEINER, T.M., PARDO, E.V., SANTOS, C.S.G., CARVALHO, A.C., WAGNER, M.F.R., GARRAFONI, A.S., BRASIL, A.C.S., RIBEIRO, Z., NOGUEIRA, J.M.M., ABBUD, A., ROSSI, M. & FUKUDA, M. 2004. Filo Annelida Classe Polychaeta. In Biodiversidade bêntica da região sul-sudeste da costa brasileira. REVIZEE Score Sul Bentos. (Amaral, A.C.Z. & C.L.D.B. Rossi-Wongtschowski, eds) São Paulo: Ulhôa Cintra Ed. p. 114-125.
- Amaral, A.C.Z. & Nonato, E.F. 1996. Annelida Polychaeta: características, glossário e chaves para famílias e gêneros da costa brasileira. Editora da Unicamp, UNICAMP, Campinas.
- AMARAL, A.C.Z., NALLIN, S.A.H. & STEINER, T.M. 2006a. Catálogo das espécies dos Annelida Polychaeta do Brasil. http://www.ib.unicamp.br/destaques/biota/bentos_marinho/prod_cien/texto_poli.pdf
- AMARAL, A.C.Z., RIZZO, A.E. & ARRUDA, E.P. (Orgs.) 2006b. Manual de Identificação dos Invertebrados Marinhos da região sudeste-sul do Brasil. Vol. I. São Paulo: Edusp Ed. 287p.
- AUGENER, H. 1934. VIII Polychaeten aus den zoologischen Museen von Leiden und Amsterdam. IV. (Schluss). Zoologische Mededeelingen uitgegeven door's Rijks Museum van Natuurlijke Historie te Leiden, 17(1-2):67-160.
- BÖGGEMANN, M. 2002. Revision of the Glyceridae Grube 1850 (Annelida: Polychaeta). Abh. Senckenberg. Naturforsch. Ges. 555:1-249.
- Böggemann, M. & Fiege, D. 2001. Description of seven new species of the genus Glycera Savigny, 1818 (Annelida: Polychaeta: Glyceridae). Ophelia 54(1):29-49.
- EHLERS, E. 1868. Die Borstenwürmer (Annelida Chaetopoda) nach systematischen und anatomischen Untersuchungen dargestellt. Verlag von Wilhelm Engelmann, Erster Band, Leipzig. p. 1-748.
- EHLERS, E. 1887. Reports on the results of dredging, under the direction of L.F. Pourtalès, during the years 1868-1870, and of Alexander Agassiz, in the Gulf of Mexico (1877-78), and in the Caribbean Sea (1878-79), in the U.S. Coast Survey Steamer Blake", Lieut.Com. C.D. Sigsbee, U.S.N., and Commander J.R. Bartlett, U.S.N., Commanding. XXXI. Report on the Annelids. Memoirs of the Museum of Comparative Zoölogy at Harvard College, Cambridge, Massachussets, v.15, p. 1-335.
- FAUCHALD, K. 1977. Polychaetes from intertidal areas in Panama, with a review of previous shallow-water records. Smith. Contr. Zool. 221:1-81.
- GILBERT, K. 1984. Family Glyceridae Grube, 1850. In Taxonomic guide to the polychaetes of the northern Gulf of Mexico (J.M. Uebelacker & P.G. Johnson). Vittor & Associates, Inc. Mobile, Alabama, p. 32-1 to 32-26.
- GRUBE, E. 1857. Annulata Örstediana. Enumeratio Annulatorum, quae in itinere per Indiam occidentalem et Americam centralem annis 1845-1848 suscepto legit cl. A.S. Örsted, adjectis specibus nonnullis a cl. H. Kröyero in itinere ad Americam meridionalem collectis (Fortsaetelse). Videnskabelige Meddelelser fra den naturhistoriske Forening I Kjöbenhavn, 158-186, Kjöbenhavn.
- GRUBE, E. 1863. Beschreibung neuer oder wenig bekannter Anneliden. Archiv für Naturgeschichte, Jahrgang 29,1:37-69, pls. 4-6, Berlin.
- GRUBE, E. 1870. Bemerkungen über die Familie der Glycereen. Jahres-Bericht der Schlesischen Gesellschaft für vaterländische Cultur, 47:56-68, Breslau.
- HARTMAN, O. 1938. Descriptions of new species and new generic records of polychaetous annelids from California of the families Glyceridae, Eunicidae, Stauronereididae and Opheliidae. Univ. Calif. Pub. Zool., 43:93-112.
- HARTMAN, O. 1968. Atlas of the Errantiate Polychaetous Annelids From California. Allan Hancock Foundation. University of Southern California, Los Angeles, Caliofrnia, 828p.
- LANA, P.C. 1984. Anelídeos poliquetas errantes do litoral do Estado do Paraná. Tese de Doutorado, Universidade Federal do Paraná, Paraná. 275p.
- LEIDY, J. 1855. Contributions towards a knowledge of the Marine Invertebrata Fauna, of the coasts of Rhode Island and New Jersey. Jornal of the Academy of Natural Sicences of Philadelphis, 3(2):135-152, plates 10-11:Philadelphia.
- NONATO, E. F. 1981. Contribuição ao Conhecimento dos Anelídeos Poliquetas Bentônicos da Plataforma Continental Brasileira, entre Cabo Frio e o Arroio Chui. Tese de Doutorado, Universidade de São Paulo, São Paulo, 246p.
- NONATO, E. & LUNA, J.A.C. 1970. Anelídeos Poliquetas do nordeste do Brasil. I - Poliquetas bentônicos da costa de Alagoas e Sergipe. Bol. Inst. Oceanogr. S. Paulo 19:57-130.
- O'CONNOR, B.D.S. 1987. The Glyceridae (Polychaeta) of the North Atlantic and Mediterranean, with descriptions of two new species. J. Nat. Hist. 21(1):167-189.
- ORENSANZ, J.M. & GIANUCA, N.M. 1974. Contribuição ao conhecimento dos anelídeos poliquetas do Rio Grande do Sul, Brazil. I. Lista sistemática preliminar e descrição de três novas espécies. Com. Mus. Ci. PUCRGS 4:1-37.
- PARRA, S., RODRÍGUEZ, C.V., LÓPEZ-JAMAR, E. & O'CONNOR, B.D.S. 1995. Contribución al conocimiento del género Glycera Savigny, 1818 (Polychaeta: Glyceridae) en la península Ibérica. Clave taxonómica y nuevos datos sobre Glycera oxycephala Ehlers, 1887. Bol. Inst. Esp. Oceanogr., 11(1):53-59.
- Quatrefages, A. de 1866. Histoire naturelle des Annelés marins et d'eau douce. Annélides et Géphyriens. Librairie Encyclopédique de Roret, 3 vols., and Atlas with pls. 1-20, Paris.
- RIZZO, A.E. & STEINER, T.M 2006. Família Glyceridae. In: Manual de Identificação dos Invertebrados Marinhos da Região Sudeste-sul do Brasil. 1 ed. São Paulo: Editora da Universidade de São Paulo, 2006, v.1:165-172.
- RULLIER, F. & AMOUREAUX, L. 1979. Annélides Polychaètes. Annales de I'Institut Océanographique Monaco 55:145-206.
- TEMPERINI, M.T. 1981. Sistemática e distribuição dos poliquetos errantes da Plataforma Continental brasileira entre as latitudes de 23° 05' S e 30° 00' S. Dissertação de Mestrado, Instituto Oceanográfico, Universidade de São Paulo, 89p.











Publication Dates
-
Publication in this collection
11 Mar 2008 -
Date of issue
2007
History
-
Accepted
01 Sept 2007 -
Reviewed
05 June 2007 -
Received
07 Feb 2007