
Abstract:
Honey pollen samples of Melipona seminigra pernigra Moure & Kerr 1950MOURE, J.S. & KERR, W.E. 1950. Sugestões para a modificação da sistemática do gênero Melipona (Hymen.-Apoidea). Dusenia 1: 105-129. sampled between 2017 and 2019 from experimental apiaries installed in campo rupestre on canga (CRC) vegetation of the Serra dos Carajás aimed to evaluated seasonal floral availability of undisturbed and mining-influenced areas. Around one hundred pollen types were identified mainly belonging to Fabaceae, Myrtaceae and Euphorbiaceae (31, 6 and 5 species, respectively). Mining area presented the highest pollen richness, almost twice those identified in the undisturbed areas. 80% of the pollen types are rare with concentrations ≤ 2,000 pollen grains/10 g, while the remaining were the most abundant, frequent and the primary bee sources. These latter correspond mostly to native plants species such as Tapirira guianensis Aubl., Protium spp., Aparisthmium cordatum (A.Juss.) Baill., Mimosa acutistipula var. ferrea Barneby, Periandra mediterranea (Vell.) Taub., Miconia spp., Pleroma carajasense K.Rocha, Myrcia splendens (Sw.) DC., Serjania spp. and Solanum crinitum Lam. All pollen types were identified during both seasons, but higher concentration values are related to the dry period (June-September). The statistical analysis of the pollen data indicated that there was no significant difference between undisturbed and mining-influenced areas, since primary bee sources of this study are widespread used in revegetation of mined areas.
Keywords:
Stingless bees; Honey; Pollen; Amazonia; Iron mining.
Resumo:
O conteúdo polínico de amostras de mel coletadas nos anos de 2017 e 2019 de apiários experimentais de Melipona seminigra pernigra Moure & Kerr 1950MOURE, J.S. & KERR, W.E. 1950. Sugestões para a modificação da sistemática do gênero Melipona (Hymen.-Apoidea). Dusenia 1: 105-129., instalado dentro de uma vegetação de campo rupestre em um afloramento de canga na Serra dos Carajás, sudeste da Amazônia, foi analisado para entender a variabilidade local dos recursos florais em áreas naturais e perturbadas. Aproximadamente 100% dos tipos polínicos foram identificados e pertencem principalmente às famílias Fabaceae, Myrtaceae e Euphorbiaceae (31, 6 e 5 espécies, respectivamente). Áreas de mineração apresentaram a maior riqueza de pólen, quase o dobro daquelas identificadas em áreas perturbadas. 80% dos tipos de pólen são raros com concentrações ≤ 2.000 grãos de pólen/10g, enquanto que os restantes foram os mais abundantes, frequentes e fontes primárias para as abelhas. Este últimos correspondem principalmente a plantas nativas como Tapirira guianensis Aubl., Protium spp., Aparisthmium cordatum (A.Juss.) Baill., Mimosa acutistipula var. ferrea Barneby, Periandra mediterrânea (Vell.) Taub., Miconia spp., Pleroma carajasense K.Rocha, Myrcia splendens (Sw.) DC., Serjania spp. e Solanum crinitum Lam. Todos os tipos polínicos foram identificados durante ambas as estações, mas altas concentrações estão relacionadas ao período seco (junho-setembro). A análise estatística indicou que não houve diferença significativa nos dados de pólen de mel entre áreas naturais e áreas anteriormente degradadas, uma vez que as fontes primárias das abelhas deste estudo são amplamente utilizadas na revegetação de áreas mineradas.
Palavras-chave:
Abelhas sem ferrão; Mel; Pólen; Amazônia; Mineração de ferro.
Introduction
Melissopalynological studies of stingless bees are important to identify the sources of nectar explored by bees and, through this, improve the management of colonies and landscape for a better honey production in both aspects: quality and quantity. Moreover, it can potentially be used to characterize the local flora regarding its diversity and phenology (Absy et al. 1984ABSY, M.L., CAMARGO, J.M.F., KERR, W.E. & MIRANDA, I.P.A. 1984. Espécies de plantas visitadas por Meliponinae (Hymenoptera; Apoidea), para coleta de pólen na região do médio Amazonas. Revista Brasileira de Biologia 44: 227-237., Absy & Kerr 1977ABSY, M.L. & KERR, W.E. 1977. Algumas plantas visitadas para obtenção de pólen por operárias de Melipona seminigra merrilae em Manaus. Acta Amazonica 7(3): 309-315.; Martins et al. 2011MARTINS, A.C.L., RÊGO, M.M.C., CARREIRA, L.M.M. & ALBUQUERQUE, P.M.C. 2011. Espectro polínico de mel de tiúba (Melipona fasciculata Smith, 1854, Hymenoptera, Apidae). Acta Amazonica 41: 183-190.). The use of this technique for identifying nectar resources used by social, native stingless bees - Meliponini tribe on Brazil dates from 1960, according to an historical review (Barth 2004BARTH, M.O. 2004. Melissopalynology in Brazil: a review of pollen analysis of honeys, propolis and pollen loads of bees. Scientia Agricola 61: 342-350.; Freitas & Novais 2014FREITAS, W.A.T. & NOVAIS, J.S. 2014. Melissopalynology in the Brazilian amazon: a databank of pollen types cited in the literature. Boletín de la Asociación Latinoamericana de Paleobotánica y Palinología 14: 103-136.; Souza et al. 2018SOUZA, R.R., ABREU, W.H.R. & NOVAIS, J.S. 2018. Melissopalynology in Brazil: a map of pollen types and published productions between 2005 and 2017. Palynology 43(4): 690-700.). Some of them have been developed specifically in the Amazon region, in natural and disturbed areas, to identify the origin of pollen in loads carried by forager bees or in the stored honey and beebread (Absy et al. 1980, Marques-Souza 1996MARQUES-SOUZA, A.C. 1996. Fontes de pólen exploradas por Melipona compressipes manaosensis (Apidae: Meliponinae), abelha da Amazônia Central. Acta Amazonica 26: 77-86., Marques-Souza et al. 1996MARQUES-SOUZA, A.C., MOURA, C.O. & NELSON, B.W. 1996. Pollen collected by Trigona williana (Hymenoptera: Apidae) in Central Amazônia. Revista de Biologia Tropical 44: 567-573., Rezende et al. 2019REZENDE, A.C.C., ABSY, M.L., FERREIRA, M.G., MARINHO, H.A. & SANTOS, O.A. 2019. Pollen of honey from Melipona seminigra merrillae Cockerell, 1919, Scaptotrigona nigrohirta Moure, 1968 and Scaptotrigona sp. Moure, 1942 (Apidae: Meliponini) reared in Satare Mawé indigenous communities, Amazon, Brazil. Palynology 43: 255-267.). These are important contributions to understand the floral resources collected by bees, especially considering the high diversity of stingless bees on Brazil and the particularities of each biome. It is also key to practical applications on sustainable development practices for rural communities (meliponiculture and honey production) and/or for the design of restoration strategies (Montoya et al. 2012MONTOYA, D., ROGERS, L. & MEMMOTT, J. 2012. Emerging perspectives in the restoration of biodiversity-based ecosystem services. TREE 27: 666-672.). Some melissopalynological studies on humid evergreen tropical forest (HETF) areas of this region have shown strong preferences of Meliponini species by Tapirira guianensis Aubl. (Anacardiaceae), Protium heptaphyllum (Aubl.) Marchand (Burseraceae) and Mimosa pudica L. (Fabaceae) (Online Resource 1; Absy & Kerr 1977; Oliveira et al. 2009OLIVEIRA, F.M., ABSY, M.L. & MIRANDA, I.S. 2009. Recurso polínico coletado por abelhas sem ferrão (Apidae, Meliponinae) em um fragmento de floresta na região de Manaus, Amazonas. Acta Amazonica 39: 505-518.; Rezende et al. 2019). These taxa widespread occur on most of Brazilian phytogeographic domains such as Amazon Rainforest, Caatinga, Central Brazilian Savanna and Atlantic Rainforest (BFG 2015BFG. 2015. Growing knowledge: an overview of Seed Plant diversity in Brazil. Rodriguésia 66(4): 1085-1113. ). As a consequence, the distribution of Meliponini species tend to follow the same domains (Camargo & Pedro 2013CAMARGO, J.M.F. & PEDRO, S.R.M. 2013. Meliponini Lepeletier, 1836. In J.S. Moure, D. Urban & G.A.R. Melo (Eds.), Catalogue of Bees (Hymenoptera, Apoidea) in the Neotropical Region - Online Version.).
Knowledge regarding bee flora in plant endemism hotspots (Kasecker et al. 2009KASECKER, T.P., SILVA, J.M.C., RAPINI, A., BARROSO, M., GIULIETTI, A., ANDRADE, M.J.G. & QUEIROZ, L. 2009. Áreas-chave para espécies raras de fanerógamas. In: Giulietti, A.M., Rapini, A., Andrade, M.J.G., Queiroz, L.P., Silva, J.M.C. (Eds.) Plantas Raras do Brasil. Conservation International, Belo Horizonte; p. 433-472.), such as campo rupestre on canga at the Serra dos Carajás (Mota et al. 2018MOTA, N.F.O., WATANABE, M.T.C., ZAPPI, D.C., HIURA, A.L., PALLOS, J., VIVEROS, R.S., GIULIETTI, A.M. & VIANA, P.L. 2018. Amazon canga: the unique vegetation of Carajás revealed by the list of seed plants. Rodriguésia 69: 1435-1487.), are still incipient (Barth 1987BARTH, O.M. 1987. Importância apícola dos campos naturais e rupestres da Serra Norte de Carajás (Pará) Boletim do Museu Parense Emílio Goeldi - Série Botânica 3(1): 21-28.). This particular type of open vegetation consists mostly of herbs and shrubs adapted to the edaphic conditions provided by iron-rich outcrops (Mota et al. 2018) surrounded mostly by evergreen tropical forest. Some Amazonian stingless bees are also poorly known, as is the case with Melipona seminigra pernigraMoure & Kerr 1950MOURE, J.S. & KERR, W.E. 1950. Sugestões para a modificação da sistemática do gênero Melipona (Hymen.-Apoidea). Dusenia 1: 105-129.. This native, abundant species in eastern Amazonia (Moure & Kerr 1950) is considered as one of the most popular species reared by traditional populations for honey production (Nogueira-Neto 1997NOGUEIRA-NETO, P. 1997. Vida e criação de abelhas indígenas sem ferrão. São Paulo (SP): Editora Nogueirapis.; Cortopassi-Laurino et al. 2006CORTOPASSI-LAURINO, M., IMPERATRIZ-FONSECA, V.L., ROUBIK, D.W., DOLLIN, A., HEARD, T., AGUILAR, I., VENTURIERI, G.C., EARDLEY, C. & NOGUEIRA-NETO, P. 2006. Global meliponiculture: challenges and opportunities. Apidologie 37: 275-292.), as seen also in the Carajás region.
Our aim is to understand the local variability of floral resources utilized by M. seminigra pernigra in natural and disturbed areas. For this, we kept colonies of this stingless bee species in experimental meliponaries on canga, forest and disturbed areas in the region of Carajás and analyzed their honey samples through melissopalynology. This is an important step for the conservation of bee species and their landscape, providing knowledge about plant species used for honey production and also to help on degraded land restoration strategies in the Carajás region.
1. Study area
The Carajás National Forest, a Federal Conservation Unit created in February 1998 comprises an area of approximately 3,930 km2 (Figure 1a). Represented by discontinuous table-top outcrops surrounded by a matrix of humid evergreen tropical forest (HETF) and seasonally dry forest (SDF). The higher areas (600-800 m a.s.l.) are occupied by lateritic plateaus with campo rupestre on canga (CRC) vegetation (Mota et al. 2018MOTA, N.F.O., WATANABE, M.T.C., ZAPPI, D.C., HIURA, A.L., PALLOS, J., VIVEROS, R.S., GIULIETTI, A.M. & VIANA, P.L. 2018. Amazon canga: the unique vegetation of Carajás revealed by the list of seed plants. Rodriguésia 69: 1435-1487.). This mineral province includes one of the largest iron ore deposits worldwide (Tolbert et al. 1971TOLBERT, G.E., TREMAINE, J.W., MELCHER, G.C. & GOMES, C.B. 1971. The recently discovered Serra dos Carajas iron deposits, northern Brazil. Economic Geology 66: 985-994.) and mining operations in the region began in 1985. The group of northern plateaus are known as Serra Norte, where two main areas are being exploited, namely N4 and N5 mines.

(a) Study site inserted in the northern portion of the Carajás National Forest, southeast Amazonia (N1 to N8 are related mining areas codes). (b) Aerial photography of the Trilha da Lagoa da Mata (small area of canga vegetation surrounded by HETF and SDF. (b) Aerial photography showing the N4 and N5 mining areas surround by canga vegetation (open areas in the right side) and (c) HETF and SDF. (d-e) sampling the honey pots.
The rainfall regime is characterized by a rainy season, regionally known as Amazonian winter from November to May, followed by a dry Amazonian summer between June and October (Lopes et al. 2013LOPES, M.N.G., SOUZA, E.B. & FERREIRA, D.B.D.S. 2013. Climatologia regional da precipitação no estado do Pará. Revista Brasileira de Climatologia 12(1): 84-102.). The total annual rainfall of the Carajás region may vary between 1,545 mm and 1,863 mm during the rainy season, with the dry season amounting to values between 159 mm and 321 mm (Silva Júnior et al. 2017SILVA Jr, R.O., SOUZA, E.B.D., TAVARES, A.L., MOTA, J.A., FERREIRA, D.B.S., SOUZA-FILHO, P.W.M. & ROCHA, E.J.P.D. 2017. Three decades of reference evapotranspiration estimates for a tropical watershed in the eastern Amazon. Anais da Academia Brasileira de Ciências 89(3): 1985-2002.). The mean temperature is 27.2°C, with a minimum annual temperature of 26.6°C in January and a maximum annual temperature of 28.1°C in September (Tavares et al. 2018TAVARES, A.L., CARMO, A.M.C., SILVA Jr, R.O., SOUZA-FILHO, P.W.M., SILVA, M.S., FERREIRA, D.B.S., NASCIMENTO Jr, W.R. & DALL’AGNOL, R. 2018. Climate indicators for a watershed in the eastern amazon. Revista Brasileira de Climatologia 23: 389-410.).
Materials and Methods
Four experimental meliponaries were installed in natural (area 1: Trilha da Lagoa da Mata) and disturbed (area 2: Viveiro Florestal; area 3: revegetation of degraded areas - RDA; area 4: N5 mine) areas of Serra Norte (Figure 1a). The area 1 is located in the northeast portion of the N5 mine, Serra Norte. It presents a narrow CRC area of around 33,000 m2 surrounded by HETF (Figure 1b). The area 2 is a plant nursery area for growing species of the Carajás flora to provide plants for the revegetation of degraded areas (RDA), and it is located ~ 1.2 km southeast of area 1, covering an area of 26,500 m2 surrounded by HETF and weedy plants; area 3 is close to the N5 mine and located 8.7 km southwest of area 1, and it consists of HETF and SDF towards the southern part of this area (Figure 1c); area 4 is a pit slope in RDA process mainly covered by CRC and weedy plants, and it is located 5.5 km southwest of area 1 and 3.4 km distant of area 3. The main plant species in the four areas are given in Online Resource 2 (Vale-Golder Associates 2011VALE-GOLDER ASSOCIATES. 2011. Projeto Área Mínima de Canga - Similaridade das paisagens de savana metalófila - 3ª aproximação (RT-020_099-515-5006_00-J). Technical Report. 444 p.), updated according Viana et al. (2016)VIANA, P.L., MOTA, N.F.O., GIL, A.S.B., SALINO, A., ZAPPI, D.C., HARLEY, R.M., ILKIU-BORGES, A.L., SECCO, R.S., ALMEIDA, T.E., WATANABE, M.T.C., DOS SANTOS, J.U.M., TROVÓ, M., MAURITY, C., GIULIETTI, A.M. 2016. Flora of the cangas of the Serra dos Carajás, Pará, Brazil: history, study area and methodology. Rodriguesia 67(5): 1107-1124.. Melipona seminigra forage range is around 1 km (L. Costa, personal communication), a flight ability similar to that of Melipona mandacaia Smith (Kuhn-Neto et al. 2009KUHN-NETO, B., CONTRERA, F.A.L., CASTRO, M.S. & NIEH, J.C. 2009. Long distance foraging and recruitment by a stingless bee, Melipona mandacaia. Apidologie 40: 472-480.), so it is unlikely that foraging activity overlapped between areas, with exception to areas 1 and 2.
Due to the expansion dynamics of the N5 mine, the meliponary installed in this area (3) was moved in March 2018 to another location in the same area, while the RDA apiary (4) started in June 2018. Monthly samples were collected from the honey pots of fourteen hives of Melipona seminigra pernigra from each apiary. The content, from five to ten honey pots (chosen randomly) was removed during each collection event (Figure 1d, e). 46 samples covering a period of 16 months (September 2017 to January 2019) were harvested.
Honey samples weighing 10 g were diluted using 20 ml of distilled water according to Maurizio & Louveaux et al. (1965)MAURIZIO, A. & LOUVEAUX, J. 1965. Pollens de plantes melliferes d’Europe. Paris: UGAF.. For each sample, a tablet containing 9,666 spores of Lycopodium clavatum was added (Stockmarr 1971STOCKMARR, J. 1971. Tablets with spores used in absolute pollen analysis. Pollen et Spores 13: 615-621.). These samples were acetolysed following Erdtman (1952)ERDTMAN, G. 1952. Pollen Morphology and Plant Taxonomy. Angiosperms. Stockholm: Almqvist and Wiksell, p. 539., and mounted on microscope slides using glycerin jelly. They were deposited in the pollen library at Instituto Tecnológico Vale (PaliITV, Belém, Brazil). Five hundred pollen grains were counted per sample using a Zeiss Axio Imager M2. Pollen type identification was based on morphological comparisons with PaliITV.
The distribution frequency of the pollen types followed Feller-Demalsy et al. (1987)FELLER-DEMALSY, M.J., PARENT, J. & STRACHAN, A.A. 1987. Microscopic analysis of honeys from Alberta, Canada. Journal of Apicultural Research 26(2): 123-132.: very frequent (>50%), frequent (20-50%), infrequent (10-20%) and rare (<10%); and groups of pollen concentration according to Maurizio (1975)MAURIZIO, A. 1975. Microscopy of honey. In: Crane, E., (Ed). Honey, a comprehensive survey.: Group I (<20,000), Group II (20,000-100,000), Group III (100,000-500,000), Group IV (500,000-1,000,000) and Group V (>1,000,000) were adopted to classify pollen types per honey sample. The range of the pollen types per honey sample was calculated using the Shannon-Weaver diversity index (H´; Ludwig and Reynolds 1988LUDWIG, J.A. & REYNOLDS, J.F. 1988. Statistical ecology: A primer on methods and computing. New York: Wiley.). Pielou’s evenness index (J’), was calculated to evaluate the degree of heterogeneity (0) or homogeneity (1) of pollen resources (Pielou 1977PIELOU, E.C. 1977. Mathematical ecology. New York, NY: Wiley.). Product moment correlation coefficient (PMCC) was applied between pollen richness, H’ and J’ data.
For statistical analysis, the total pollen concentration (grains/cm³ unit) of four areas (Trilha da Lagoa da Mata, Viveiro Florestal, N5 Mine and RDA) were counted based on the definition of the seasonal rainfall pattern. The descriptive analyses were performed using common and abundant taxa among sites, and Wilcoxon test for significance difference of each site between periods. Bray-Curtis based non-metric multidimensional scaling (NMDS) has been applied to evaluate species distribution and their relationship between the sites. The NMDS analysis required the vegan R package (version 2.5.3; Oksanen 2016OKSANEN, J., BLANCHET, F.G., FRIENDLY, M., KINDT, R., LEGENDRE, P., MCGLINN, D., MINCHIN, P.R., O’HARA, R.B., SIMPSON, G.L., SOLYMOS, P., STEVENS, M.H.H., SZOECS, E. & WAGNER, H. 2016. vegan: Community Ecology Package. R package version 2.4-1. https://CRAN.R-project.org/package=vegan
https://CRAN.R-project.org/package=vegan...
), and visualization were carried out using the ggplot2 package (version 3.4.1; Wickham 2009WICKHAM, H. 2009. ggplot2: Elegant Graphics for Data Analysis. New York: Springer-Verlag.)
Climatological data covering the collection years were obtained from the meteorological station of Serra Norte de Carajás (OMM: 81860), which covers all studied areas.
Results
1. Climatological data
The patterns of wind intensity and direction in the study site from January 2017 to January 2019 show slight variation of wind speed (~ 1 to 2 m/s) with prevailing direction from the E and ENE (Figure 1a). The mean minimum and maximum air temperature for this period was 23.8 ºC and 27.7 ºC, respectively, with the highest values in August to September, while the lowest temperatures occurred in February and July (Figure 2a). Based on the accumulated monthly rainfall (Figure 2a), the dry period extended from June to September, ranging from 67 to 132 mm. The rainy period comprised October to May, with values ranging from 1805 to 2300 mm. The total annual precipitation during 2017 was lower than that for 2018 with values of 1935 and 2367 mm, respectively. The wettest month of the study period was February 2019 (approx. 700 mm) Relative humidity follows the seasonal rainfall pattern, with the lowest values (< 80%) observed during in the dry period. Highest solar irradiance was detected in July with a mean of ~ 220 W/m2, while the lowest occurred in December, with ~ 140 W/m2 (Figure 2b).

(a) Accumulated monthly rainfall data (mm; right side) and mean air temperature (ºC; left side) at the Carajás Meteorological Station from January 2017 to January 2019. (b) Monthly mean solar irradiance (W/m2; right side) and Maximum and minimum relative humidity (%; left side). Gray bands indicate the drier climate periods.
2. Pollen assemblage
A total of 104 pollen types were identified in the 46 honey samples analyzed, belonging to 35 families. Fabaceae, Myrtaceae and Euphorbiaceae were the most representative with 31, 6 and 5 species, respectively (Online Resource 3).
In general, the N5 mine presented the highest pollen richness with 95 pollen types identified. Trilha da Lagoa da Mata and Viveiro Florestal presented around 60 pollen types each, while RDA had 39 types. Maximum pollen concentration per area ranged from 60,895 to 1,521,194 pollen grains/g, which were observed, respectively, in RDA and Viveiro Florestal areas. 80% of the pollen types were found with values < 5,000 grains/10g and < 1%. Thus, the pollen types are generally rare (< 10%; Feller-Demalsy et al. 1987FELLER-DEMALSY, M.J., PARENT, J. & STRACHAN, A.A. 1987. Microscopic analysis of honeys from Alberta, Canada. Journal of Apicultural Research 26(2): 123-132.) and belong to group I (≤ 20,000 pollen grains/10 g; Maurizio, 1975MAURIZIO, A. 1975. Microscopy of honey. In: Crane, E., (Ed). Honey, a comprehensive survey.) (see Online Resource 3; Online Resource 4).
The remaining 20% of the pollen types are the most abundant and present in all areas along the studied period (Online Resource 3; Online Resource 5, Online Resource 6, Online Resource 7, Online Resource 8). These types were identified as Tapirira guianensis (Anacardiaceae), Protium (Burseraceae), Aparisthmium cordatum and Alchornea (Euphorbiaceae), Mimosa acutistipula var. ferrea and Periandra mediterranea (Fabaceae), Miconia and Pleroma carajasense (Melastomataceae), Myrcia splendens (Myrtaceae), Serjania (Sapindaceae) and Solanum crinitum (Solanaceae) (Figure 3, Figure 4) abundances varied according to the year of study.

Optical microscopy photomicrographs of the most abundant and frequent pollen types identified in this study. Tapirira guianensis (1, 2); Protium (3, 4), Aparisthmium cordatum (5, 6) and Alchornea (7, 8), Mimosa acutistipula var. ferrea (9, 10) and Periandra mediterranea (11-14), Miconia (17, 18) and Pleroma carajasense (19, 20). Scale bars: 10 μm.

Figure 4. Optical microscopy photomicrographs of the most abundant and frequent pollen types identified in this study. Myrcia splendens (1-4), Serjania (5-8) and Solanum crinitum (9-12). Scale bars: 10 μm.
Significantly higher pollen concentration was detected on 2018 (Online Resource 5, Online Resource 6, Online Resource 7, Online Resource 8). Thus, for the Trilha da Lagoa da Mata, pollen types of P. carajasense, Miconia, Alchornea and M. acutistipula var. ferrea abounded in March and April, while Protium and T. guianensis in August and September (Online Resource 5).
Protium and T. guianensis also abounded in June and July in the Viveiro Florestal, where A. cordatum was very representative in October to December (Online Resource 6). Protium was abundant in June and November in the RDA area (Online Resource 8).
The pollen types M. splendens and M. acutistipula var. ferrea were significantly present from September to December 2018, but not during the same period of the previous year in N5 mine.
According to the mean values of pollen concentration of the most abundant pollen types during the dry and rainy seasons, the pollen of all plant species was identified during both seasons, however in variable abundance. Higher values are generally related to the dry period (June-September). M. acutistipula var. ferrea is more abundant at the Viveiro Florestal and N5 mine in the dry and wet season, respectively. P. carajasense abounded during the wet season in the Trilha da Lagoa da Mata, while it was remarkably important in the dry season in the Viveiro Florestal (Figure 5).

Total pollen concentration of the most abundant, frequent and common plant species of all studied areas considering the different climate periods.
The Shannon-Weaver diversity (H’) and Pielou’s evenness (J’) indexes have strong PMCC (ρ-value ~ 0.7 to 0.9), and moderate to strong correlation with pollen richness (ρ-value ~ 0.5 to 0.9) in each area (Figure 6). Thus, higher H’ and J’ were related to 2017, while lower values were mainly found during June and July 2018.

Richness (gray bars), Shannon-Weaver diversity (H’; upper stright line with empty circles) and Pielou’s evenness (J’; lower dashed line with black ircles) of the pollen data per sampling area: (a) Lagoa da Trilha, (b) Viveiro Florestal, (c) Mina N5 and (d) RDA areas.
3. Statistical analyses
Considering the statistical analysis, the overall mean of pollen concentration of all sites shows an overlap of pollen concentration distribution (Figure 7a). RDA samples had the lowest concentration than all other sites for both periods. Diversely, Viveiro Florestal had different abundance for the two periods. Wilcoxon test was applied to evaluate differences between climate periods for each site. The results show no statistically significant difference on sites between periods, except for the Viveiro Florestal (ρ-value = 0.03). In order to evaluate the pollen distribution and similarity among the sites, Non-metric Multidimensional Scaling was the appropriate ordination method because the stress value for two dimensional, which was 0.04. NMDS, pointed to (dis)similarities on sites based on pollen concentration (Figure 7b). RDA samples are more related on both periods due to abundance of the pollen types of Protium and Serjania. N5 Mine samples are more similar during both periods due to the abundance of Mimosa acutistipula var. ferrea and Myrcia splendens, whereas the Trilha da Lagoa da Mata and Viveiro Florestal (rainy period) have more contribution from A. cordatum, Miconia and P. carajasense. During the dry period, Trilha da Lagoa da Mata and Viveiro Florestal differed from other samples due to their pollen assemblages: the first is mainly composed by T. guianensis and Protium and the latter has higher influence from several species. According to the Wilcoxon test, Viveiro Florestal demonstrated significant statistical difference for both periods, and therefore the NMDS is coherent with Wilcoxon test. Viveiro Florestal had higher concentration of all pollen assemblages during the dry period, whereas, during the rainy period it has a composition more comparable to the one at the Trilha da Lagoa da Mata and N5 Mine in the same period.
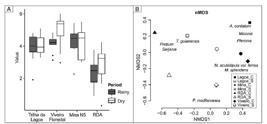
(a) Boxplot of pairwise samples in rainy and dry period. (b) Result of Non-Metric Multidimensional Scaling using honey samples in two distinct periods (dark gray: rainy period; white: dry period). Named pollen taxa used in the analysis are included.
Discussion
Considering the total pollen assemblage identified in honey samples of M. seminigra pernigra, around 80% of the pollen types were found to be within the values < 5,000 grains/10g and < 1%. The remaining ~20% are the primary bee sources. The secondary sources (less abundant) are continuously explored, providing a small amount of food that is an alternative resource for the colony (Heinrich 1976HEINRICH, B. 1976. The foraging specializations of individual bumblebees. Ecological Monographs 46: 105-128.). This is especially important when other plants are saturated by other pollinators or flowering diminishes according to seasonal changes. In some occasions, the secondary source moves to a central position in the food supply (Novais & Absy 2013NOVAIS, J.S. & ABSY, M.L. 2013. Palynological examination of the pollen pots of native stingless bees from the Lower Amazon region in Pará, Brazil. Palynology 37(2): 218-230.).
Melipona species are a successful generalist group of bees from tropical humid forests that have a close relationship with tree crowns and mass flowering (Ramalho 2004RAMALHO, M. 2004. Stingless bees and mass flowering trees in the canopy of Atlantic Forest: a tight relationship. Acta Botanica Brasilica 18(1): 37-47.), and consequently play an important role in plant reproduction and natural forest regeneration. In the present experimental study developed at the Carajás National Forest, the honey samples of M. seminigra pernigra were constituted mostly by native species pollen.
Considered a native and abundant species, Mimosa acutistipula var. ferrea Barneby is endemic to north and northeastern Brazil and abundant in the Serra dos Carajás region, occurring on all CRC outcrops studied (Mattos et al. 2018MATTOS, C.M.J., SILVA e SILVA, W.L., CARVALHO, C.S., LIMA, A.N., FARIA, S.M. & LIMA, H.C. 2018. Flora das cangas da serra dos Carajás, Pará, Brasil: Leguminosae. Rodriguésia 69(3): 1147-1220.). It features in the Brazilian red list of plants as Data Deficient (DD) (Martinelli & Moraes 2013MARTINELLI, G. & MORAES, M.N. 2013. Livro vermelho da flora do Brasil. Jardim Botânico do Rio de Janeiro, Rio de Janeiro.) and vulnerable (VU) in the List of Threatened Species of Pará State (COEMA-PA 2010COEMA - CONSELHO ESTADUAL DE MEIO AMBIENTE, PARÁ. 2010. Instrução Normativa n. 02/2010, Diário Oficial do Estado do Pará, 2010.). M. acutistipula var. ferrea forms large single-species stands in both canga and altered barren areas (Zappi et al. 2018ZAPPI, D.C., GASTAUER, M., RAMOS, S., NUNES, S., CALDEIRA, C.F., SOUZA-FILHO, P.W., GUIMARÃES, J.T.F., GIANNINI, T.C., VIANA, V.P.L., LOVO, J., MOTA, N.F.O., SIQUEIRA, J.O. 2018. Plantas nativas para recuperação de áreas de mineração em Carajás. Belém, PA: Instituto Tecnológico Vale (ITV). 282 p.). The clustered, long, pendulous, sweetly scented inflorescences of M. acutistipula var. ferrea are very rich in pollen (Mattos et al. 2018). This mass flowering is very attractive for M. seminigra pernigra. The abundance of the Mimosa-type (small) pollen grains in honey samples can be related to the configuration and relatively small pollen size, varying from 10-14 µm (Zappi et al. 2018), which are also released in large amounts according Ferreira and Absy (2017)FERREIRA, M.G. & ABSY, M.L. 2017. Pollen analysis of honeys of Melipona (Michmelia) seminigra merrillae and Melipona (Melikerria) interrupta (Hymenoptera: Apidae) bred in Central Amazon, Brazil. Grana 56(6): 436-449..
P. carajasense is a recently described endemic species from the CRC of Carajás (Rocha et al. 2017ROCHA, K.C.J., GOLDENBERG, R., VIANA, P.L. & MEYER, F.S. 2017. Pleroma carajasense (Melastomataceae), a new species endemic to ironstone outcrops in the Brazilian Amazon. Phytotaxa 329(3): 233-242.), and it has been categorized as a possible Endangered (EN) species according to the IUCN criteria (IUCN 2012IUCN. 2012. IUCN Red list categories. Version 3.1, 2nd edition IUCN Species Survival Commission. Cambridge (UK): Gland and Cambridge. World Conservation Union.). It occurs in large populations both on natural and anthropic areas. This species has attractive short thyrsoid inflorescences with 7-25 campanulate flowers, lilac-pink to purple petals and poricidal anthers (Matos & Santos 2017MATOS, V.R. & SANTOS, F.A.R. 2017. Pollen in honey of Melipona scutellaris L. (Hymenoptera: Apidae) in an Atlantic rainforest area in Bahia, Brazil. Palynology 41: 144-156., Rocha et al. 2017). Vibrating bees (flower or anther buzzing) are favored by poricidal anthers, limiting the access of competing pollinators (De Luca & Vallejo‐Marín 2013DE LUCA, P.A. & VALLEJO‐MARÍN, M. 2013. What’s the “buzz” about? The ecology and evolutionary significance of buzz‐pollination. Current Opinion in Plant Biology 16: 429-435.). M. seminigra pernigra have body sizes exceeding the gap between anthers and stigma in P. carajasense, and can be considered an efficient pollinator (Solís‐Montero & Vallejo‐Marín 2017SOLÍS‐MONTERO, L. & VALLEJO‐MARÍN, M. 2017. Does the morphological fit between flowers and pollinators affect pollen deposition? An experimental test in a buzz‐pollinated species with anther dimorphism. Ecology and Evolution 7: 2706-2715.). Considering Alchornea spp., nectary stomata is commonly observed on flowers, also in leaves, of A. discolor Poepp., A. acutifolia Müll. Arg., A. castaneifolia (Willd.) A. Juss., A. glandulosa Poepp., A. megalophylla Müll. Arg., which were extensively described for the study area (R. Secco personal communication; Secco 2004).
Other abundant plant species visited by M. seminigra pernigra in the study area, but with wider geographical distribution, are A. cordatum, M. splendens and T. guianensis. These species are commonly observed in forest margins, gallery forest, seasonally dry forest (SDF) and humid evergreen tropical forest (HETF), as well as disturbed areas in the Amazon and elsewhere. A. cordatum has racemose inflorescences with female flowers closer to the foliage and distal male flowers, while the two latter are paniculate. All these inflorescences provide large number of flowers per inflorescence. All species are good source of pollen and nectar for bees, as well as resin in the case of T. guianensis (Matos & Santos 2017MATOS, V.R. & SANTOS, F.A.R. 2017. Pollen in honey of Melipona scutellaris L. (Hymenoptera: Apidae) in an Atlantic rainforest area in Bahia, Brazil. Palynology 41: 144-156.). P. mediterranea has also a wide distribution with preference for open ecosystems such as Amazonian savannas, high altitude grasslands, campo rupestre on canga and other substrates (Mattos et al. 2018MATTOS, C.M.J., SILVA e SILVA, W.L., CARVALHO, C.S., LIMA, A.N., FARIA, S.M. & LIMA, H.C. 2018. Flora das cangas da serra dos Carajás, Pará, Brasil: Leguminosae. Rodriguésia 69(3): 1147-1220.). It has racemose inflorescence with few, showy blue to purplish open, zygomorphic pea-shaped flowers with conspicuous nectar guides. These flowers provide both pollen and nectar for bees (Mateus 1998MATEUS, S. 1998. Abundância relativa, fenologia e visita às flores pelo Apoidea do cerrado da Estação Ecológica de Jataí - Luiz Antônio - SP [master’s thesis]. Universidade de São Paulo, Faculdade de Filosofia e Letras de Ribeirão Preto.).
Pollen type of Miconia is frequently abundant in honey samples of Melipona bees in the Amazon region (Absy et al. 1980ABSY, M.L., BEZERRA, E.B. & KERR, W.E. 1980. Plantas nectaríferas utilizadas por duas espécies de Melipona da Amazônia. Acta Amazonica 10: 271-281., Marques-Souza 1996MARQUES-SOUZA, A.C. 1996. Fontes de pólen exploradas por Melipona compressipes manaosensis (Apidae: Meliponinae), abelha da Amazônia Central. Acta Amazonica 26: 77-86., Oliveira et al. 2009OLIVEIRA, F.M., ABSY, M.L. & MIRANDA, I.S. 2009. Recurso polínico coletado por abelhas sem ferrão (Apidae, Meliponinae) em um fragmento de floresta na região de Manaus, Amazonas. Acta Amazonica 39: 505-518., Ferreira & Absy 2017FERREIRA, M.G. & ABSY, M.L. 2017. Pollen analysis of honeys of Melipona (Michmelia) seminigra merrillae and Melipona (Melikerria) interrupta (Hymenoptera: Apidae) bred in Central Amazon, Brazil. Grana 56(6): 436-449.). The continuous flowering along both dry and wet seasons provides an excellent source of pollen for bees (Renner 1989RENNER, S.S. 1989. A survey of reproductive biology in Neotropical Melastomataceae and Memecylaceae. Annals of the Missouri Botanical Garden. 76: 496-518.). Miconia has poricidal anthers, supporting selective buzz pollination (Buchmann 1983BUCHMANN, S.L. 1983. Buzz pollination in angiosperms. In: Jones, G.E., Little, R.J., (Eds.) Handbook of experimental pollination biology, New York: Van Nostrand Reinhold; p. 73-113.). However, the identification of Miconia to species level using honey samples was not possible due to the stenopalynous pollen found in the genus, and there are at least 20 species of Miconia cited for the CRC of Carajás (Rocha et al. 2017ROCHA, K.C.J., GOLDENBERG, R., VIANA, P.L. & MEYER, F.S. 2017. Pleroma carajasense (Melastomataceae), a new species endemic to ironstone outcrops in the Brazilian Amazon. Phytotaxa 329(3): 233-242.). This large genus, one of the biggest in the Brazilian Flora, comprises 288 species in Brazil, 81 of them recorded for Pará state (BFG 2020BFG. 2015. Growing knowledge: an overview of Seed Plant diversity in Brazil. Rodriguésia 66(4): 1085-1113. ), and its species present considerable variation in the gap between anthers and stigma. Therefore, it is not possible to be sure whether M. seminigra pernigra effectively makes contact with the stigmas.
Pollen types of Solanum crinitum were present in lower frequency in honey samples than Miconia but were well-represented in the study site. It has also poricidal anthers, but in this case the stamens are fused into a cone and the anthers are almost in contact (Giacomin & Gomes 2018GIACOMIN, L.L. & GOMES, E.S.C. 2018. Flora das cangas da Serra dos Carajás, Pará, Brasil: Solanaceae. Rodriguésia 69(3): 1373-1396.). Thus, bees can vibrate all anthers at once, performing effective pollination (Solís‐Montero & Vallejo‐Marín 2017SOLÍS‐MONTERO, L. & VALLEJO‐MARÍN, M. 2017. Does the morphological fit between flowers and pollinators affect pollen deposition? An experimental test in a buzz‐pollinated species with anther dimorphism. Ecology and Evolution 7: 2706-2715.). Melipona has been commonly observed visiting Solanaceae species in lowland Amazonian forest (Absy et al. 1980ABSY, M.L., BEZERRA, E.B. & KERR, W.E. 1980. Plantas nectaríferas utilizadas por duas espécies de Melipona da Amazônia. Acta Amazonica 10: 271-281.), and, according to these authors, its pollen is of great importance for the diet and maintenance of the Melipona hives. Extrafloral nectaries are reported for Solanum (Anderson & Symon 1985ANDERSON, G.J. & SYMON, D.E. 1985. Extrafloral nectaries in Solanum. Biotropica 17(1):40-45.). Predominant occurrence of Solanum pollen types with different sizes in the same honey sample of Melipona are also reported in the northeastern Brazil (Ferreira & Absy 2015FERREIRA, M.G. & ABSY, M.L. 2015. Pollen niche and trophic interactions between colonies of Melipona (Michmelia) seminigramerrillae and Melipona (Melikerria) interrupta (Apidae: Meliponini) reared in floodplains in the Central Amazon. Arthropod Plant Interactions 9: 263-279., 2017FERREIRA, M.G. & ABSY, M.L. 2017. Pollen analysis of honeys of Melipona (Michmelia) seminigra merrillae and Melipona (Melikerria) interrupta (Hymenoptera: Apidae) bred in Central Amazon, Brazil. Grana 56(6): 436-449.). However, it is possible that the flowering of Solanum crinitum Lam., which is abundant in the study site (Giacomin & Gomes 2018) and certainly very rich in pollen, may has concomitantly occurred with any other nectar provider species. As the pollen extraction from the anthers of Solanum is done by vibration (buzz pollination; Buchmann & Cane 1989BUCHMANN, S.L. & CANE, J.H. 1989. Bees assess pollen returns while sonicating Solanum flowers. Oecologia 81: 289-294.), the body hairs of M. seminigra pernigra may be full of pollen, which are moved to the corbicula. In the hives, the pollen is deposited into pots, but it is tasted by other workers of the colony (Roubik 1989ROUBIK, D.W. 1989. Ecology and natural history of tropical bees. 1° edn. New York, NY: Cambridge University Press; p. 514.). Likewise, the nectar collected is transferred between workers by trophallaxis before its deposition into the pots. Thus, the contamination of nectar collected from other source with the Solanum pollen that is present in the digestive system of the workers may be also a plausible explanation.
Also well represented in honey samples were pollen types of Protium and Serjania, chiefly at the Trilha da Lagoa da Mata, Viveiro Florestal and N5 mine areas. Protium is the commonest component in the pollen assemblage of sampled honey at the RDA area. These taxa present paniculate inflorescences with small, separate sex flowers that provide bees with nectar, resin and pollen (male flowers only), while Serjania spp. is a rampant liana with thyrsoid inflorescences with small, sweetly scented flowers that are considered an excellent source of nectar (Matos & Santos 2017MATOS, V.R. & SANTOS, F.A.R. 2017. Pollen in honey of Melipona scutellaris L. (Hymenoptera: Apidae) in an Atlantic rainforest area in Bahia, Brazil. Palynology 41: 144-156.). Both genera are commonly observed in transitional areas between CRC and HETF and SDF and provide great availability of resources over a long flowering period (Oliveira et al. 2009OLIVEIRA, F.M., ABSY, M.L. & MIRANDA, I.S. 2009. Recurso polínico coletado por abelhas sem ferrão (Apidae, Meliponinae) em um fragmento de floresta na região de Manaus, Amazonas. Acta Amazonica 39: 505-518.).
Following the statistical analysis, there was no significant difference in honey pollen data between the natural and disturbed areas. This is mainly due to the fact that all these abundant and common native taxa are used in revegetation during RDA processes by mining activities (Zappi et al. 2018ZAPPI, D.C., GASTAUER, M., RAMOS, S., NUNES, S., CALDEIRA, C.F., SOUZA-FILHO, P.W., GUIMARÃES, J.T.F., GIANNINI, T.C., VIANA, V.P.L., LOVO, J., MOTA, N.F.O., SIQUEIRA, J.O. 2018. Plantas nativas para recuperação de áreas de mineração em Carajás. Belém, PA: Instituto Tecnológico Vale (ITV). 282 p.). However, more studies are necessary to better evaluate the prolonged effects of human interference over plant reproduction.
Regarding the importance of plants for honey production, income generation, and conservation of pollinators, T. guianensis, Protium spp., M. splendens, M. acutistipula var. ferrea, Serjania spp., P. carajasense and A. cordatum seem to be key species, as well as Miconia spp. with some considerations. Indeed, most of Miconia species are polliniferous, but it cannot be generalized (T. Vasconcelos, personal communication). Most neotropical Melastomataceae have bee-pollinated flowers with poricidal anthers. However, nectar rewards are known to be produced in about 80 species in eight genera, including Miconia (Varassin et al. 2008VARASSIN, I., PENNEYS, D. & MICHELANGELI, F. 2008. Comparative Anatomy and Morphology of Nectar-producing Melastomataceae. Annals of Botany 102(6): 899-909., Brito et al. 2017BRITO, V.L.G., RECH, A.R., OLLERTON, J. & SAZIMA, M. 2017. Nectar production, reproductive success and the evolution of generalised pollination within a specialised pollen-rewarding plant family: a case study using Miconia theizans. Plant Systematics and Evolution 303: 709-718.). These authors using anatomical methods based on scanning electron microscopy, and serial sections of paraffin-embedded flowers precisely identified nectary stomata on the ovary apex in Miconia spp.
These plant species can be used to help recover deforested areas (Zappi et al. 2018ZAPPI, D.C., GASTAUER, M., RAMOS, S., NUNES, S., CALDEIRA, C.F., SOUZA-FILHO, P.W., GUIMARÃES, J.T.F., GIANNINI, T.C., VIANA, V.P.L., LOVO, J., MOTA, N.F.O., SIQUEIRA, J.O. 2018. Plantas nativas para recuperação de áreas de mineração em Carajás. Belém, PA: Instituto Tecnológico Vale (ITV). 282 p.) and help improve the resource provisions for the stingless bee populations. For local beekeepers, the incorporation of these plants in the foraging area of beehives can help to improve the amount of honey produced, reflecting positively on income generation.
Conclusions
M. seminigra pernigra uses, preferably, native plants from cangas and forests of the Serra do Carajás as its primary sources. Main examples are Tapirira guianensis Aubl., Protium spp., Aparisthmium cordatum (A.Juss.) Baill., Mimosa acutistipula var. ferrea Barneby, Periandra mediterrânea (Vell.) Taub., Miconia spp., Pleroma carajasense K.Rocha, Myrcia splendens (Sw.) DC., Serjania spp., and Solanum crinitum Lam. However, these primary bee sources (PBS) represent only 20% of the total pollen assemblage, which suggest high dispersion rates of the colony members related to temporal flowering pattern of the PBS. All pollen types were identified during both seasons, but higher concentration values are related to the dry period (June-September). In fact, Shannon-Weaver diversity (H') and Pielou’s evenness (J') indexes substantially decrease when PBS are fully available.
The PBS have different floral syndromes. The strategies included condensed spike-shaped inflorescences offering abundant nectar and pollen in M. acutistipula var. ferrea and A. cordatum; buzz pollinated anthers in P. carajensis, Miconia spp. and S. crinitum; and separate sex flowers (A. cordatum and Protium spp.). The majority of the species had small flowers grouped in inflorescences working as a flowering unit, while only P. mediterranea and P. carajasense have large, attractive flowers. Of these species, the only strongly zygomorphic flower with nectar guides was P. mediterranea.
No significant difference was statically found in honey pollen data between the natural and disturbed areas, which may be related to the widespread use of PBS for mine land rehabilitation, as well as in the remaining vegetation surrounding the mine. However, longer-term data are necessary for better supporting such findings in mining land areas of the study site, including climate change influences.
Acknowledgments
The authors thank the members of Meio Ambiente Corredor Norte of Vale S.A. for the field support and scientific discussions. This project was carried out in the National Forest of Carajas under the permission of IBAMA (SISBIO 35594-2), and was funded by ITV-VALE S.A. (RBRS000603.17.02.MS02). We would like to thank the associated editor and anonymous reviewers for their valuable comments.
Supplementary material
The following online material is available for this article:
Online Resource 1 - Melissopalynological records of plant/pollen associated with Meliponini species (Rezende et al. 2019REZENDE, A.C.C., ABSY, M.L., FERREIRA, M.G., MARINHO, H.A. & SANTOS, O.A. 2019. Pollen of honey from Melipona seminigra merrillae Cockerell, 1919, Scaptotrigona nigrohirta Moure, 1968 and Scaptotrigona sp. Moure, 1942 (Apidae: Meliponini) reared in Satare Mawé indigenous communities, Amazon, Brazil. Palynology 43: 255-267.; Oliveira et al. 2009OLIVEIRA, F.M., ABSY, M.L. & MIRANDA, I.S. 2009. Recurso polínico coletado por abelhas sem ferrão (Apidae, Meliponinae) em um fragmento de floresta na região de Manaus, Amazonas. Acta Amazonica 39: 505-518.; Absy & Kerr 1977ABSY, M.L. & KERR, W.E. 1977. Algumas plantas visitadas para obtenção de pólen por operárias de Melipona seminigra merrilae em Manaus. Acta Amazonica 7(3): 309-315.; Marques-Souza et al. 2002MARQUES-SOUZA, A.C., MIRANDA, I.P.A., MOURA, C.O., RABELO, A., BARBOSA, E.M. 2002. Características morfológicas e bioquímicas do pólen coletado por cinco espécies de Meliponíneos da Amazônia Central. Acta Amazonica 32(2): 217-229.; Ferreira & Absy 2017.
Online Resource 2 - Main plant species observed in the natural and anthropized areas were the studied apiaries were installed. Humid Evergreen Tropical Forest (HETF), Seasonally Dry Forest (SDF).
Online Resource 3 - Occurrence per area and distribution frequency of the pollen types (Feller-Demalsy et al., 1987FELLER-DEMALSY, M.J., PARENT, J. & STRACHAN, A.A. 1987. Microscopic analysis of honeys from Alberta, Canada. Journal of Apicultural Research 26(2): 123-132.): very frequent (>50%), frequent (freq.: 20–50%), infrequent (infreq.: 10–20%) and rare (<10%); and groups of pollen concentration (Maurizio, 1975MAURIZIO, A. 1975. Microscopy of honey. In: Crane, E., (Ed). Honey, a comprehensive survey.): Group I (<20,000), Group II (20,000–100,000), Group III (100,000–500,000), Group IV (500,000–1,000,000) and Group V (>1,000,000).
Online Resource 4 - Number of observations per (1) groups of pollen concentration (grains/10g; Maurizio, 1975MAURIZIO, A. 1975. Microscopy of honey. In: Crane, E., (Ed). Honey, a comprehensive survey.): Group I (<20,000), Group II (20,000–100,000), Group III (100,000–500,000), Group IV (500,000–1,000,000) and Group V (>1,000,000); and per (2) distribution frequency of the pollen types (percent-%; Feller-Demalsy et al., 1987): very frequent (>50), frequent (20–50), infrequent (10–20) and rare (<10). https://minio.scielo.br/documentstore/1676-0611/WY3pxQHf5QBvXmskmBTYH6z/ddc614ac686108c77949e2c995d8d1de2f615dda.pdf
Online Resource 5 - Percentage and concentration pollen diagram illustrating the pollen content of the honey samples from Trilha da Lagoa da Mata area. Gray line indicates a gap in the honey sampling. https://minio.scielo.br/documentstore/1676-0611/WY3pxQHf5QBvXmskmBTYH6z/94cee3f7e43a406e4c090067a6db3faed3abc71a.pdf
Online Resource 6 - Percentage and concentration pollen diagram illustrating the pollen content of the honey samples from Viveiro Florestal area. Gray lines indicate gaps in the honey sampling. https://minio.scielo.br/documentstore/1676-0611/WY3pxQHf5QBvXmskmBTYH6z/e41ca382d9f9aa2722ea708e526665407662c6c7.pdf
Online Resource 7 - Percentage and concentration pollen diagram illustrating the pollen content of the honey samples from N5 mine. Gray line indicates a gap in the honey sampling. https://minio.scielo.br/documentstore/1676-0611/WY3pxQHf5QBvXmskmBTYH6z/c5f205f713a20573c37754c6458ff642308c2d6b.pdf
Online Resource 8 - Percentage and concentration pollen diagram illustrating the pollen content of the honey samples from RDA area. https://minio.scielo.br/documentstore/1676-0611/WY3pxQHf5QBvXmskmBTYH6z/bc725cceeeb20a4d5a89faa5096f729c3ee300cc.pdf
References
- ABSY, M.L. & KERR, W.E. 1977. Algumas plantas visitadas para obtenção de pólen por operárias de Melipona seminigra merrilae em Manaus. Acta Amazonica 7(3): 309-315.
- ABSY, M.L., BEZERRA, E.B. & KERR, W.E. 1980. Plantas nectaríferas utilizadas por duas espécies de Melipona da Amazônia. Acta Amazonica 10: 271-281.
- ABSY, M.L., CAMARGO, J.M.F., KERR, W.E. & MIRANDA, I.P.A. 1984. Espécies de plantas visitadas por Meliponinae (Hymenoptera; Apoidea), para coleta de pólen na região do médio Amazonas. Revista Brasileira de Biologia 44: 227-237.
- ANDERSON, G.J. & SYMON, D.E. 1985. Extrafloral nectaries in Solanum Biotropica 17(1):40-45.
- BARTH, O.M. 1987. Importância apícola dos campos naturais e rupestres da Serra Norte de Carajás (Pará) Boletim do Museu Parense Emílio Goeldi - Série Botânica 3(1): 21-28.
- BARTH, M.O. 2004. Melissopalynology in Brazil: a review of pollen analysis of honeys, propolis and pollen loads of bees. Scientia Agricola 61: 342-350.
- BFG. 2015. Growing knowledge: an overview of Seed Plant diversity in Brazil. Rodriguésia 66(4): 1085-1113.
- BRITO, V.L.G., RECH, A.R., OLLERTON, J. & SAZIMA, M. 2017. Nectar production, reproductive success and the evolution of generalised pollination within a specialised pollen-rewarding plant family: a case study using Miconia theizans. Plant Systematics and Evolution 303: 709-718.
- BUCHMANN, S.L. & CANE, J.H. 1989. Bees assess pollen returns while sonicating Solanum flowers. Oecologia 81: 289-294.
- BUCHMANN, S.L. 1983. Buzz pollination in angiosperms. In: Jones, G.E., Little, R.J., (Eds.) Handbook of experimental pollination biology, New York: Van Nostrand Reinhold; p. 73-113.
- CAMARGO, J.M.F. & PEDRO, S.R.M. 2013. Meliponini Lepeletier, 1836. In J.S. Moure, D. Urban & G.A.R. Melo (Eds.), Catalogue of Bees (Hymenoptera, Apoidea) in the Neotropical Region - Online Version.
- COEMA - CONSELHO ESTADUAL DE MEIO AMBIENTE, PARÁ. 2010. Instrução Normativa n. 02/2010, Diário Oficial do Estado do Pará, 2010.
- CORTOPASSI-LAURINO, M., IMPERATRIZ-FONSECA, V.L., ROUBIK, D.W., DOLLIN, A., HEARD, T., AGUILAR, I., VENTURIERI, G.C., EARDLEY, C. & NOGUEIRA-NETO, P. 2006. Global meliponiculture: challenges and opportunities. Apidologie 37: 275-292.
- DE LUCA, P.A. & VALLEJO‐MARÍN, M. 2013. What’s the “buzz” about? The ecology and evolutionary significance of buzz‐pollination. Current Opinion in Plant Biology 16: 429-435.
- ERDTMAN, G. 1952. Pollen Morphology and Plant Taxonomy. Angiosperms. Stockholm: Almqvist and Wiksell, p. 539.
- FELLER-DEMALSY, M.J., PARENT, J. & STRACHAN, A.A. 1987. Microscopic analysis of honeys from Alberta, Canada. Journal of Apicultural Research 26(2): 123-132.
- FERREIRA, M.G. & ABSY, M.L. 2015. Pollen niche and trophic interactions between colonies of Melipona (Michmelia) seminigramerrillae and Melipona (Melikerria) interrupta (Apidae: Meliponini) reared in floodplains in the Central Amazon. Arthropod Plant Interactions 9: 263-279.
- FERREIRA, M.G. & ABSY, M.L. 2017. Pollen analysis of honeys of Melipona (Michmelia) seminigra merrillae and Melipona (Melikerria) interrupta (Hymenoptera: Apidae) bred in Central Amazon, Brazil. Grana 56(6): 436-449.
- FREITAS, W.A.T. & NOVAIS, J.S. 2014. Melissopalynology in the Brazilian amazon: a databank of pollen types cited in the literature. Boletín de la Asociación Latinoamericana de Paleobotánica y Palinología 14: 103-136.
- GIACOMIN, L.L. & GOMES, E.S.C. 2018. Flora das cangas da Serra dos Carajás, Pará, Brasil: Solanaceae. Rodriguésia 69(3): 1373-1396.
- GUIMARÃES, J.T.F., COSTA, L., ZAPPI, D.C., BATISTA JUNIOR, W.F., LOPES, K.D.S., ALVES, R.C.D.O., ROMEIRO, L.D.A., SILVA, E.F.D., CARREIRA, L.M.M., RODRIGUES, T.M., GIANNINI, T.C., IMPERATRIZ-FONSECA, V.L., BARTH, O.M. 2019. Melissopalynology Reveals the Foraging Preferences of the Stingless Bee Melipona seminigra pernigra Moure & Kerr 1950 (Apidae: Meliponini) in Cangas of Serra dos Carajás, Southeastern Amazonia. Preprints, 2019100134 (doi: 10.20944/preprints201910.0134.v1)
» https://doi.org/10.20944/preprints201910.0134.v1 - GUIMARÃES, J.T.F., GIANNINI, T.C., VIANA, V.P.L., LOVO, J., MOTA, N.F.O. & SIQUEIRA, J.O. 2018. Plantas nativas para recuperação de áreas de mineração em Carajás Belém, PA: Instituto Tecnológico Vale (ITV), p. 282.
- HEINRICH, B. 1976. The foraging specializations of individual bumblebees. Ecological Monographs 46: 105-128.
- IUCN. 2012. IUCN Red list categories. Version 3.1, 2nd edition IUCN Species Survival Commission. Cambridge (UK): Gland and Cambridge. World Conservation Union.
- JAFFÉ, R., POPE, N., CARVALHO, A.T., MAIA, U.M., BLOCHTEIN, B., CARVALHO, C.A.L., CARVALHO-ZILSE, G.A., FREITAS, B.M., MENEZES, C., RIBEIRO, M.F., VENTURIERI, G.C. & IMPERATRIZ-FONSECA, V.L. 2015. Bees for development: Brazilian survey reveals how to optimize stingless beekeeping. Plos One 10(6): e0130111.
- KUHN-NETO, B., CONTRERA, F.A.L., CASTRO, M.S. & NIEH, J.C. 2009. Long distance foraging and recruitment by a stingless bee, Melipona mandacaia Apidologie 40: 472-480.
- KASECKER, T.P., SILVA, J.M.C., RAPINI, A., BARROSO, M., GIULIETTI, A., ANDRADE, M.J.G. & QUEIROZ, L. 2009. Áreas-chave para espécies raras de fanerógamas. In: Giulietti, A.M., Rapini, A., Andrade, M.J.G., Queiroz, L.P., Silva, J.M.C. (Eds.) Plantas Raras do Brasil. Conservation International, Belo Horizonte; p. 433-472.
- LOPES, M.N.G., SOUZA, E.B. & FERREIRA, D.B.D.S. 2013. Climatologia regional da precipitação no estado do Pará. Revista Brasileira de Climatologia 12(1): 84-102.
- LOUVEAUX, J., MAURIZIO, A. & VORWOHL, G. (1978). Methods of melissopalynology. Bee World 59: 139-157.
- LUDWIG, J.A. & REYNOLDS, J.F. 1988. Statistical ecology: A primer on methods and computing. New York: Wiley.
- MARTINELLI, G. & MORAES, M.N. 2013. Livro vermelho da flora do Brasil. Jardim Botânico do Rio de Janeiro, Rio de Janeiro.
- MARQUES-SOUZA, A.C. 1996. Fontes de pólen exploradas por Melipona compressipes manaosensis (Apidae: Meliponinae), abelha da Amazônia Central. Acta Amazonica 26: 77-86.
- MARQUES-SOUZA, A.C., MOURA, C.O. & NELSON, B.W. 1996. Pollen collected by Trigona williana (Hymenoptera: Apidae) in Central Amazônia. Revista de Biologia Tropical 44: 567-573.
- MARQUES-SOUZA, A.C., MIRANDA, I.P.A., MOURA, C.O., RABELO, A., BARBOSA, E.M. 2002. Características morfológicas e bioquímicas do pólen coletado por cinco espécies de Meliponíneos da Amazônia Central. Acta Amazonica 32(2): 217-229.
- MARTINS, A.C.L., RÊGO, M.M.C., CARREIRA, L.M.M. & ALBUQUERQUE, P.M.C. 2011. Espectro polínico de mel de tiúba (Melipona fasciculata Smith, 1854, Hymenoptera, Apidae). Acta Amazonica 41: 183-190.
- MATEUS, S. 1998. Abundância relativa, fenologia e visita às flores pelo Apoidea do cerrado da Estação Ecológica de Jataí - Luiz Antônio - SP [master’s thesis]. Universidade de São Paulo, Faculdade de Filosofia e Letras de Ribeirão Preto.
- MATOS, V.R. & SANTOS, F.A.R. 2017. Pollen in honey of Melipona scutellaris L. (Hymenoptera: Apidae) in an Atlantic rainforest area in Bahia, Brazil. Palynology 41: 144-156.
- MATTOS, C.M.J., SILVA e SILVA, W.L., CARVALHO, C.S., LIMA, A.N., FARIA, S.M. & LIMA, H.C. 2018. Flora das cangas da serra dos Carajás, Pará, Brasil: Leguminosae. Rodriguésia 69(3): 1147-1220.
- MAURIZIO, A. & LOUVEAUX, J. 1965. Pollens de plantes melliferes d’Europe. Paris: UGAF.
- MAURIZIO, A. 1975. Microscopy of honey. In: Crane, E., (Ed). Honey, a comprehensive survey.
- MONTOYA, D., ROGERS, L. & MEMMOTT, J. 2012. Emerging perspectives in the restoration of biodiversity-based ecosystem services. TREE 27: 666-672.
- MOTA, N.F.O., WATANABE, M.T.C., ZAPPI, D.C., HIURA, A.L., PALLOS, J., VIVEROS, R.S., GIULIETTI, A.M. & VIANA, P.L. 2018. Amazon canga: the unique vegetation of Carajás revealed by the list of seed plants. Rodriguésia 69: 1435-1487.
- MOURE, J.S. & KERR, W.E. 1950. Sugestões para a modificação da sistemática do gênero Melipona (Hymen.-Apoidea). Dusenia 1: 105-129.
- NOGUEIRA-NETO, P. 1997. Vida e criação de abelhas indígenas sem ferrão. São Paulo (SP): Editora Nogueirapis.
- NOVAIS, J.S. & ABSY, M.L. 2013. Palynological examination of the pollen pots of native stingless bees from the Lower Amazon region in Pará, Brazil. Palynology 37(2): 218-230.
- OKSANEN, J., BLANCHET, F.G., FRIENDLY, M., KINDT, R., LEGENDRE, P., MCGLINN, D., MINCHIN, P.R., O’HARA, R.B., SIMPSON, G.L., SOLYMOS, P., STEVENS, M.H.H., SZOECS, E. & WAGNER, H. 2016. vegan: Community Ecology Package. R package version 2.4-1. https://CRAN.R-project.org/package=vegan
» https://CRAN.R-project.org/package=vegan - OLIVEIRA, F.M., ABSY, M.L. & MIRANDA, I.S. 2009. Recurso polínico coletado por abelhas sem ferrão (Apidae, Meliponinae) em um fragmento de floresta na região de Manaus, Amazonas. Acta Amazonica 39: 505-518.
- PIELOU, E.C. 1977. Mathematical ecology. New York, NY: Wiley.
- QUEZADA-EUÁN, J.J.G., NATES-PARRA, G., MAUÉS, M.M., IMPERATRIZ-FONSECA, V.L. & ROUBIK, D.W. 2018. Economic and Cultural Values of Stingless Bees (Hymenoptera: Meliponini) among Ethnic Groups of Tropical America. Sociobiology 65: 534-557.
- RAMALHO, M. 2004. Stingless bees and mass flowering trees in the canopy of Atlantic Forest: a tight relationship. Acta Botanica Brasilica 18(1): 37-47.
- RENNER, S.S. 1989. A survey of reproductive biology in Neotropical Melastomataceae and Memecylaceae. Annals of the Missouri Botanical Garden. 76: 496-518.
- REZENDE, A.C.C., ABSY, M.L., FERREIRA, M.G., MARINHO, H.A. & SANTOS, O.A. 2019. Pollen of honey from Melipona seminigra merrillae Cockerell, 1919, Scaptotrigona nigrohirta Moure, 1968 and Scaptotrigona sp. Moure, 1942 (Apidae: Meliponini) reared in Satare Mawé indigenous communities, Amazon, Brazil. Palynology 43: 255-267.
- ROCHA, K.C.J., GOLDENBERG, R., VIANA, P.L. & MEYER, F.S. 2017. Pleroma carajasense (Melastomataceae), a new species endemic to ironstone outcrops in the Brazilian Amazon. Phytotaxa 329(3): 233-242.
- ROUBIK, D.W. 1989. Ecology and natural history of tropical bees 1° edn. New York, NY: Cambridge University Press; p. 514.
- SILVA Jr, R.O., SOUZA, E.B.D., TAVARES, A.L., MOTA, J.A., FERREIRA, D.B.S., SOUZA-FILHO, P.W.M. & ROCHA, E.J.P.D. 2017. Three decades of reference evapotranspiration estimates for a tropical watershed in the eastern Amazon. Anais da Academia Brasileira de Ciências 89(3): 1985-2002.
- SOUZA, R.R., ABREU, W.H.R. & NOVAIS, J.S. 2018. Melissopalynology in Brazil: a map of pollen types and published productions between 2005 and 2017. Palynology 43(4): 690-700.
- SOLÍS‐MONTERO, L. & VALLEJO‐MARÍN, M. 2017. Does the morphological fit between flowers and pollinators affect pollen deposition? An experimental test in a buzz‐pollinated species with anther dimorphism. Ecology and Evolution 7: 2706-2715.
- SOSA-NÁJERA, M.S., MARTÍNEZ-HENÁNDEZ, E., LOZANO-GARCÍA, M.S. & CUADRIELLO-AGUIAR, J.I. 1994. Nectaropolliniferous sources used by Trigona (Tetragonisca) angustula in Chiapas, Southern México. Grana 33: 225-230.
- STOCKMARR, J. 1971. Tablets with spores used in absolute pollen analysis. Pollen et Spores 13: 615-621.
- TAVARES, A.L., CARMO, A.M.C., SILVA Jr, R.O., SOUZA-FILHO, P.W.M., SILVA, M.S., FERREIRA, D.B.S., NASCIMENTO Jr, W.R. & DALL’AGNOL, R. 2018. Climate indicators for a watershed in the eastern amazon. Revista Brasileira de Climatologia 23: 389-410.
- TOLBERT, G.E., TREMAINE, J.W., MELCHER, G.C. & GOMES, C.B. 1971. The recently discovered Serra dos Carajas iron deposits, northern Brazil. Economic Geology 66: 985-994.
- VALE-GOLDER ASSOCIATES. 2011. Projeto Área Mínima de Canga - Similaridade das paisagens de savana metalófila - 3ª aproximação (RT-020_099-515-5006_00-J). Technical Report. 444 p.
- VARASSIN, I., PENNEYS, D. & MICHELANGELI, F. 2008. Comparative Anatomy and Morphology of Nectar-producing Melastomataceae. Annals of Botany 102(6): 899-909.
- VIANA, P.L., MOTA, N.F.O., GIL, A.S.B., SALINO, A., ZAPPI, D.C., HARLEY, R.M., ILKIU-BORGES, A.L., SECCO, R.S., ALMEIDA, T.E., WATANABE, M.T.C., DOS SANTOS, J.U.M., TROVÓ, M., MAURITY, C., GIULIETTI, A.M. 2016. Flora of the cangas of the Serra dos Carajás, Pará, Brazil: history, study area and methodology. Rodriguesia 67(5): 1107-1124.
- WICKHAM, H. 2009. ggplot2: Elegant Graphics for Data Analysis. New York: Springer-Verlag.
- ZAPPI, D.C., GASTAUER, M., RAMOS, S., NUNES, S., CALDEIRA, C.F., SOUZA-FILHO, P.W., GUIMARÃES, J.T.F., GIANNINI, T.C., VIANA, V.P.L., LOVO, J., MOTA, N.F.O., SIQUEIRA, J.O. 2018. Plantas nativas para recuperação de áreas de mineração em Carajás. Belém, PA: Instituto Tecnológico Vale (ITV). 282 p.
Publication Dates
-
Publication in this collection
03 Mar 2021 -
Date of issue
2021
History
-
Received
02 Apr 2020 -
Reviewed
13 Oct 2020 -
Accepted
06 Dec 2020







